Файл: 1. Понятие об иммунитете и иммунологии. Предмет, задачи и основные иммунологии. Иммунология immunis.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 09.11.2023
Просмотров: 199
Скачиваний: 2
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
В соответствии с Международной классификацией антитела называются иммуноглобулинами и обозначаются Ig.
Иммуноглобулины делят на классы, а также на подклассы. Известно 5 классов: IgG, IgM, IgA, IgD, IgE.
Иммуноглобулины – это белки, построенные из нескольких полипептидных цепей. Молекула каждого класса состоит из 4 полипептидных цепей – двух тяжелых и двух легких, которые связанны между собой дисульфидными мостиками. Мелкие цепи (I) – общие для всех классов и подклассов. Тяжелые цепи (Н) имеют характерные особенности строения у каждого класса и подкласса.
При своей специфичности антитела неоднородны и отличаются друг от друга, они гетерогенны. Существует более 100000 антигенов и к каждому из них синтезируется «свое» специфическое антитело. Антитела реагируют с антигенами благодаря наличию у них определенных структур – активных центров. Активный центр представляет собой полость или щель, которая по конфигурации соответствует детерминантной группе антигена. Активный центр, куда входит детерминантная группа, должен быть ей комплиментарен, без этого не поступит феномен серологической специфичности.
Молекулы антител различных классов различают по валентности, т.е. по количеству у них активных центров. Так, IgG и IgA бивалентны (обладают двумя активными центрами), IgM поливалентен, может связать 5 – 10 молекул антигена.
Активность связывания антител с антигеном оценивается такими понятиями, как аффинитет и авидность. Аффинитет характеризует степень совпадения
(комплементарности) конфигураций активного центра антитела и антигенной детерминанты (как ключ входит в замочную скважину). Под авидностью понимают количество (валентность) и расположение активных центров, характеризующие «жадность» связывания с антигеном всей молекулы антитела.
Функции антител в организме:
1. Взаимодействие с комплементарными структурами антигена с целью его нейтрализации и последующей элиминации.
2. Обеспечение кооперации иммунокомпетентных клеток.
3. Участвуют в различных защитных реакциях организма (активации комплемента, фагоцитозе – опсонизирующее действие, аллергии, иммунологической памяти и толерантности).
4. Антитоксический эффект.
5. Цитотоксический эффект.
6. Иммунорегулирующие свойства.
7. Образование циркулирующих иммунных комплексов (ЦИК)
Иммуноглобулины класса G (IgG) – основной иммуноглобулин сыворотки здорового человека, составляет 70–75 % всей фракции иммуноглобулинов.
IgG наиболее активны во вторичном иммунном ответе и антитоксическом иммунитете. Являются единственной фракцией иммуноглобулинов, способной к транспорту через плацентарный барьер и тем самым обеспечивают иммунитет плода и новорожденного. IgG являются самыми небольшими молекулами по сравнению с другими классами Ig (молекулярная масса М = 146 кДа).
Иммуноглобулины класса А (IgA) содержатся в сыворотке (15–20 % всей фракции Ig), в секреторном компоненте: слюне, слезах, молозиве, молоке, отделяемом слизистой оболочки мочеполовой и респираторной системы. IgA представлены в виде мономеров (80 % в сыворотке), димеров (в секрете), и тримеров. Средняя молекулярная масса IgA М = 500 кДа.
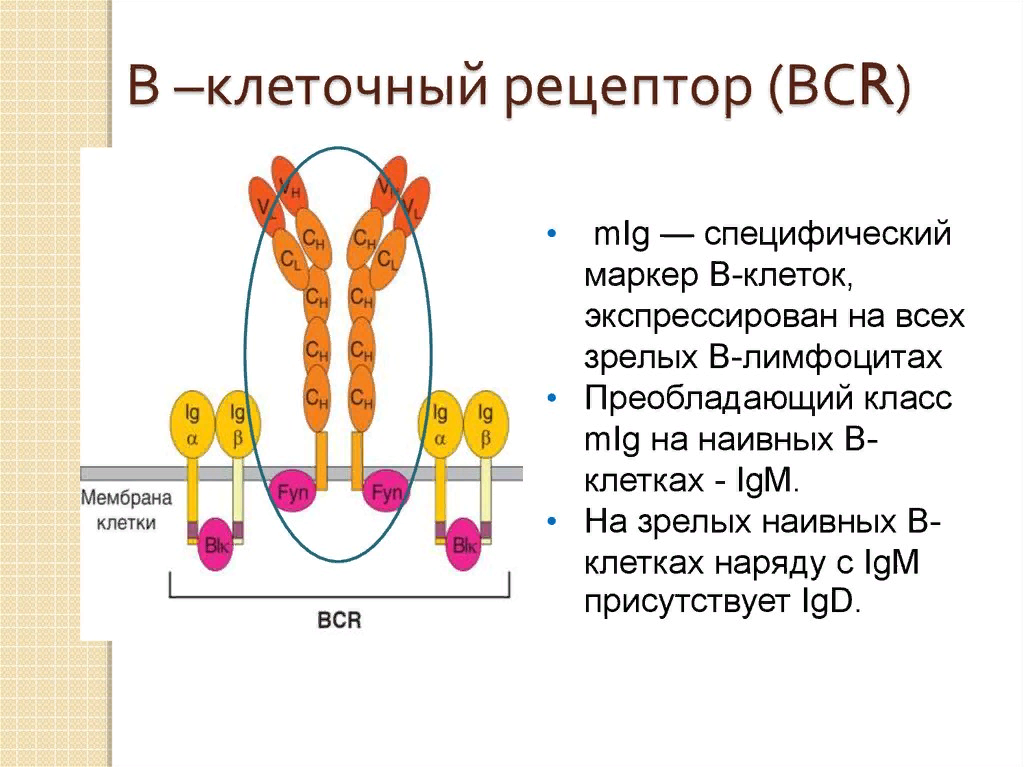
Иммуноглобулины класса М (IgM) являются пентамерами IgG и составляют до 10 % фракции иммуноглобулинов. Появляются IgM при первичном иммунном ответе B-лимфоцитами на неизвестный антиген. IgM встроены в плазматическую мембрану B-лимфоцитов и выполняют роль антиген распознающего рецептора. Средняя молекулярная масса IgМ М = 970 кДа.
Иммуноглобулины класса D (IgD) представлены в виде мономеров и содержатся в основном на мембране некоторых В-лимфоцитов, а также в небольшом количестве в плазме (< 1 % Ig плазмы). Функции IgD до конца не выяснены, предположительно они являются антигенными рецепторами с высоким содержанием связанных с белком углеводов для В-лимфоцитов, еще не представлявшихся антигену. М = 175 кДа.
Иммуноглобулины класса Е (IgE) – мономеры. Их функция заключается в связывании с поверхностью базофилов и тучных клеток, с последующим присоединением к ним антигена, при этом происходит дегрануляцию и выброс в кровь гистамина и серотонина – медиаторов воспаления. IgE участвуют в защите от паразитарных инфекций, обуславливают многие аллергические реакции. М = 200 кДа.
Антитела также классифицируют по типу антигена и, соответственно, по выполняемым Ат функциям. По этому принципу можно выделить несколько основных классов антител:
● Антиинфекционные или антипаразитарные антитела, вызывающие гибель или нарушение жизнедеятельности возбудителя инфекции либо паразита.
● Антитоксические антитела, не вызывающие гибели самого возбудителя или паразита, но обезвреживающие вырабатываемые им токсины, либо другие экзогенные токсические вещества.
● «Антитела-свидетели заболевания», наличие которых в организме сигнализирует о знакомстве иммунной системы с данным антигеном
(возбудителем) в прошлом или о текущем присутствии этого антигена
(возбудителя), но которые не играют существенной роли в борьбе организма с возбудителем (не обезвреживают ни самого возбудителя, ни его токсины, а связываются со второстепенными белками возбудителя).
● Естественные антитела – антитела к эндогенным соединениям, присутствующие в здоровом организме и выполняющие регуляторные функции;
● Аутоагрессивные антитела, или аутологичные антитела, аутоантитела – антитела, способные образовываться из естественных антител и вызывающие разрушение или повреждение нормальных, здоровых тканей самого организма хозяина и запускающие механизм развития аутоиммунных заболеваний.
● Аллореактивные антитела, или гомологичные антитела, аллоантитела – антитела против антигенов тканей или клеток других организмов того же биологического вида. Аллоантитела играют важную роль в процессах отторжения аллотрансплантантов, например, при пересадке почки, печени, костного мозга, и в реакциях на переливание несовместимой крови.
● Гетерологичные антитела, или изоантитела – антитела против антигенов тканей или клеток организмов других биологических видов. Изоантитела являются причиной невозможности осуществления ксенотрансплантации даже между эволюционно близкими видами (например, невозможна пересадка печени шимпанзе человеку) или видами, имеющими близкие иммунологические и антигенные характеристики (невозможна пересадка органов свиньи человеку). К изоантителам относятся также вторичные
(антивидовые) антитела, используемые в различных видах ИХМ.
● Антиидиотипические антитела – антитела против антител, вырабатываемых самим же организмом
17. Структура и функции различных классов иммуноглобулинов
18. Методы изучения гуморального иммунитета. (преципитация, агглютинация, реакция связывания комплемента, биологическая нейтрализация, иммобилизация, антитело-зависимая цитотоксичность, иммунофлюоресцентный анализ, иммуноферментный анализ, радиоиммунопреципитация).
19. Строение и функционирование В-клеточного рецептора.
20. Первичный и вторичный иммунный ответ. Метод парных сывороток.
В ПЕРВИЧНОМ ИММУННОМ ОТВЕТЕ гуморального типа выделяют несколько этапов. ПЕРВЫЙ
ЭТАП —латентная фаза, длится до 4 суток с момента внедрения антигена. (В этот период происходит
фагоцитоз антигена, его переработка и представление Т- и В- лимфоцитам, кооперативные
взаимодействия клеток участвующих в иммунном ответе).
ВТОРОЙ ЭТАП (лог-фаза) - синтез антител и логарифмическое удвоение их титра. На этом
этапе образовавшиеся плазматические клетки осуществляют синтез специфических антител, причем
их титр удваивается каждые 2-4 ч. по мере включения клеток клона в иммунный ответ. К 10 - 12
суткам титр антител достигает максимума. 3 ЭТАП стабилизация синтеза антител. Этот период
характеризуется динамическим равновесием между продукцией иммуноглобулинов и их элиминацией в составе иммунных комплексов. При элиминации антигена наступает 4 ЭТАП —снижение титра антител.
При первичном иммунном ответе в основном синтезируются антитела, относящиеся к Ig M E.
Лишь в конце первичного иммунного ответа происходит синтез антител класса Ig G.
При ВТОРИЧНОМ ИММУННОМ ОТВЕТЕ, при повторном контакте с антигеном, первыми в
иммунный ответ вступают клетки памяти. При вторичном иммунном ответе латентный период и
период логарифмического увеличения титра антител сокращаются примерно вдвое. В основном
синтезируются Ig G E, максимальный уровень которых превышает максимальный уровень антител
при первичном иммунном ответе.

21. Основные этапы дифференцировки Т-лимфоцитов в тимусе. Рецепторный аппарат. Виды Т-лимфоцитов.
22. Позитивная и негативная селекция. Миграция и расселение Тлимфоцитов в организме.
Положительный отбор. Дважды-положительные (DN) тимоциты (CD4+CD8+) мигрируют глубоко в кору тимуса, где они представлены самоантигенами. Эти собственные антигены (аутоантигены) экспрессируются эпителиальными клетками коры тимуса на MHC-молекулы на поверхности кортикальных (корковых) эпителиальных клеток. Только те тимоциты, которые взаимодействуют с MHC-I или MHC-II, получат жизненно важный «сигнал выживания». Те, которые недостаточно сильно взаимодействуют - погибнут без сигнала выживания. Этот процесс гарантирует, что выбранные Т-клетки будут обладать сродством к MHC, которое может выполнять полезные функции в организме (т.е. клетки должны иметь возможность взаимодействовать с MHC и пептидными комплексами, чтобы вызывать иммунные ответы). Подавляющее большинство развивающихся тимоцитов погибнет во время этого процесса. Процесс положительного отбора занимает несколько дней.
Судьба тимоцита определяется в ходе положительного отбора. Двойные положительные клетки (CD4+CD8+), которые хорошо взаимодействуют с молекулами MHC класса II, в конечном итоге станут CD4+ клетками, в то время как тимоциты, которые хорошо взаимодействуют с молекулами MHC класса I, созревают в CD8+ клетки. Клетка становится CD4+ клеткой, за счет подавления экспрессии своих поверхностных CD8-рецепторов. Если клетка не потеряет свой сигнал, она продолжит снижать регуляцию CD8 и станет CD4+, единственной положительной клеткой.
Этот процесс не удаляет тимоциты, которые могут вызывать аутоиммунитет. Потенциально аутоиммунные клетки удаляются в процессе отрицательного отбора, который происходит в мозговом веществе тимуса.
Отрицательный отбор. Отрицательный отбор удаляет тимоциты, способные прочно связываться с «собственными» пептидами MHC. Тимоциты, пережившие положительный отбор, мигрируют к границе коры и продолговатого мозга в тимусе. Находясь в продолговатом мозге, они снова представлены самоантигеном, представленным комплексом МНС медуллярных эпителиальных клеток тимуса (mTECs). mTECs должны быть AIRE+, чтобы правильно экспрессировать аутоантигены из всех тканей организма на их пептидах MHC класса I. Некоторые mTECs фагоцитозируются тимическими дендритными клетками; это позволяет представлять собственные антигены на молекулах MHC класса II (положительно отобранные CD4+ клетки должны взаимодействовать с молекулами MHC класса II, поэтому APCs, обладающие MHC класса II, должны присутствовать для отрицательного отбора CD4+ Т-клеток). Тимоциты, которые слишком сильно взаимодействуют с самоантигеном, получают апоптопический сигнал, который приводит к гибели клеток. Однако некоторые из этих клеток выбираются для превращения в Treg-клетки. Оставшиеся клетки выходят из тимуса в виде зрелых наивных Т-клеток. Этот процесс является важным компонентом центральной толерантности и служит для предотвращения образования самореактивных Т-клеток, способных индуцировать аутоиммунные заболевания у хозяина.
Прим. ред.: Наивная Т-клетка представляет собой Т-клетку, которая дифференцировалась в тимусе и успешно прошла положительный и отрицательный процессы центральной селекции в тимусе. Среди них наивные формы Т-хелперов (CD4+) и цитотоксические Т-клетки (CD8+). Наивные Т-клетки считаются незрелыми и, в отличие от активированных Т-клеток или Т-клеток памяти, не сталкивались со своим родственным антигеном на периферии.
23. Структура и функционирование Т-клеточного рецептора (TCR). Корецепторные молекулы.
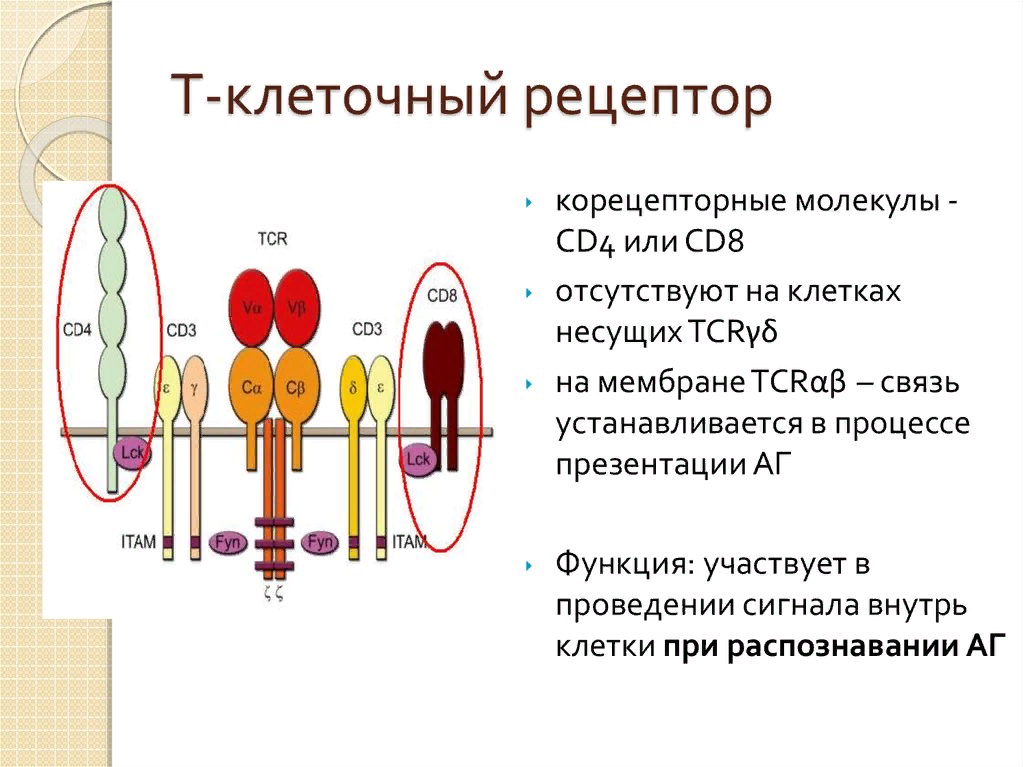
24. Распознавание антигена, выбор CD4- или CD8-пути Т-клеточного ответа.
В процессе участвуют также Т-клеточные корецепторные молекулы:
-
CD8 – для взаимодействия c молекулами MHC-I
-
CD4 – для взаимодействия c молекулами MHC-II
25. Механизм специфической цитотоксичности. Физиологическое и патологическое значение Т-клеточной цитотоксичности.
26. Методы изучения Т-клеточного звена. Методы изучения клеточной цитотоксичности.
27. Общие представления о системе комплемента. История открытия.
Комплементом называют сложный комплекс белков, действующий совместно для удаления внеклеточных форм патогена; система активируется спонтанно определенными патогенами или комплексом антиген:антитело. Активированные белки либо непосредственно разрушают патоген (киллерное действие), либо обеспечивают лучшее их поглощение фагоцитами (опсонизирующее действие); либо выполняют функцию хемотаксических факторов, привлекая в зону проникновения патогена клетки воспаления.
Комплекс белков комплемента формирует каскадные системы, обнаруженные в плазме крови. Для этих систем характерно формирование быстрого, многократно усиленного ответа на первичный сигнал за счет каскадного процесса. В этом случае продукт одной реакции служит катализатором последующей, что в конечном итоге приводит к лизису клетки или микроорганизма.
Существует два главных пути (механизма) активации комплемента - классический и альтернативный.
Классический путь активации комплемента инициируется взаимодействием компонента комплемента С1q с иммунными комплексами ( антителами, связанными с поверхностными антигенами бактериальной клетки); в результате последующего развития каскада реакций образуются белки с цитолитической (киллерной) активностью, опсонины, хемоаттрактанты. Такой механизм соединяет приобретенный иммунитет (антитела) с врожденным иммунитетом (комплемент).
Альтернативный путь активации комплемента инициируется взаимодействием компонента комплемента С3b с поверхностью бактериальной клетки; активация происходит без участия антител. Данный путь активации комплемента относится к факторам врожденного иммунитета.
В целом система комплемента относится к основным системам врожденного иммунитета, функция которых состоит в том, чтобы отличить "свое" от "не своего". Эта дифференциация в системе комплемента осуществляется благодаря присутствию на собственных клетках организма регуляторных молекул, подавляющих активацию комплемента.
C-белки системы комплемента
Все белки классического пути активации комплемента и белки лизирующего мембрану комплекса имеют буквенное обозначение "C" и арабскую цифру, отражающую последовательность открытия белка, но не последовательность его включения в реакцию. Реакционная последовательность представляет следующий ряд:
C1 , C1q , C1r , C1s , C4
Заметим, что при расщеплении компонентов системы комплемента большему продукту присваивается символ "b", а меньшему - символ "a". Из этого правила есть одно исключение:C2b означает меньший, а C2a - больший фрагмент.
Среди этих белков них много предшественников ферментов - проферментов, которые приобретают активность только после расщепления. Обозначение активного фермента отличается от обозначения его неактивного предшественника обычно надбуквенной чертой, а в данном обзоре - волнистой, например: C1r
.
CD8 – для взаимодействия c молекулами MHC-I
CD4 – для взаимодействия c молекулами MHC-II
Белок C1 построен из 5 молекул: одной C1q , двух C1r и двух C1s ( рис. 9.22 ). Пары молекул C1r и C1s располагаются поперек молекулы C1q. Соединение этих молекул зависит от ионов кальция.
С1q состоит из шести идентичных субъединиц, конформационно напоминающих булаву с коллагеноподобной рукоятью.
Взаимодействие С1q с иммунными комплексами приводит к активации комплемента.
C1s содержит участки аминокислотной последовательности из сериновой эстеразы и рецептора липопротеинов низкой плотности, а также короткий общий повтор, встречающийся в суперсемействе регуляторных белков комплемента.
Наиболее важным компонентом системы комплемента является C3 , присутствующий в плазме крови в той же концентрации (1-2 мг/мл), что и некоторые иммуноглобулины.
C3- это бетаглобулин с мол. массой равной 195 кД, секретируется (как про-С3) макрофагами.
C3 постоянно расщепляется на C3а и C3b . Внутренняя тиоэфирная связь в нативной молекуле C3 чувствительна к спонтанному гидролизу. Эта постоянная, происходящая на низком уровне самопроизвольная активация C3 в плазме называется "холостой", и она поддерживает в плазме крови небольшую концентрацию C3b.
Расщепление C3 на C3a и C3b ( C3-конвертазой в процессе активации системы) является центральным моментом любого из каскадов комплемента.
C4 (C4-bp - binding protein) - гептамерный белок плазмы, молекула которого имеет паукообразную форму; относится к суперсемейству регуляторных белков комплемента.
Каскад всех реакций активации комплемента завершается образованием лизирующего (атакующего) мембрану комплекса (ЛМК).
Первый этап образования комплекса - это ферментативное расщепление C5-белка комплемента. Белок C5 гомологичен белкам C3 и C4, но не содержит внутренней тиоэфирной связи. Прежде чем подвергнуться расщеплению C5-конвертазой, белок C5 избирательно связывается с C3b , который находится в составе конвертазы.
При расщеплении C5 высвобождаются небольшой фрагмент C5a и фрагмент C5b . C5a является высокоактивным анафилатоксином. С фрагмента C5b начинается следующий этап формирования ЛМК.
C6 , C7 , C8 и C9 - компоненты лизирующего мембрану комплекса, свойства которых аналогичны перфорину цитотоксических T-клеток и катионному белку эозинофилов
.
В конце XIX столетия было установлено, что сыворотка крови содержит некий «фактор», обладающий бактерицидными свойствами. В 1896 году молодой бельгийский ученый Жюль Борде, работавший в Институте Пастера в Париже, показал, что в сыворотке имеются два разных вещества, совместное действие которых приводит к лизису бактерий: термостабильный фактор и термолабильный (теряющий свои свойства при нагревании сыворотки) фактор. Термостабильный фактор, как оказалось, мог действовать только против определенных микроорганизмов, в то время как термолабильный фактор имел неспецифическую антибактериальную активность. Термолабильный фактор позднее был названкомплементом. Термин «комплемент» ввел Пауль Эрлих в конце 1890-х годов. Эрлих был автором гуморальной теории иммунитета и ввел в иммунологию много терминов, которые впоследствии стали общепринятыми. Согласно его теории, клетки, ответственные за иммунные реакции, имеют на поверхности рецепторы, которые служат для распознаванияантигенов. Эти рецепторы мы сейчас называем «антителами» (основой вариабельного рецептора лимфоцитов является прикреплённое к мембране антитело класса IgD, реже IgM. Антитела других классов в отсутствие соответствующего антигена не прикреплены к клеткам). Рецепторы связываются с определенным антигеном, а также с термолабильным антибактериальным компонентом сыворотки крови. Эрлих назвал термолабильный фактор «комплементом» потому, что этот компонент крови «служит дополнением» к клеткам иммунной системы.
Эрлих полагал, что имеется множество комплементов, каждый из которых связывается со своим рецептором, подобно тому, как рецептор связывается со специфическим антигеном. В противоположность этому Борде утверждал, что существует «дополнение» только одного типа. В начале XX века спор был разрешен в пользу Борде; выяснилось, что комплемент может активироваться с участием специфических антител или самостоятельно, неспецифическим способом.
Общее представление Компоненты системы комплемента
Комплемент — система белков, включающая около 20 взаимодействующих компонентов: С1 (комплекс из трех белков), С2, СЗ, …, С9, фактор В, фактор D и ряд регуляторных белков. Все эти компоненты — растворимые белки с мол. массой от 24 000 до 400 000, циркулирующие в крови и тканевой жидкости. Белки комплемента синтезируются в основном в печени и составляют приблизительно 5 % от всей глобулиновой фракции плазмы крови. Большинство из них неактивны до тех пор, пока не будут приведены в действие или в результате иммунного ответа (с участием антител), или непосредственно внедрившимся микроорганизмом (см. ниже). Один из возможных результатов активации комплемента — последовательное объединение так называемых поздних компонентов (С5, С6, С7, С8 и С9) в большой белковый комплекс, вызывающий лизис клеток (литический, или мембраноатакующий, комплекс). Агрегация поздних компонентов происходит в результате ряда последовательных реакций протеолитической активации с участием ранних компонентов (С1, С2, С3, С4, фактора В и фактора D). Большинство этих ранних компонентов — проферменты, последовательно активируемые путем протеолиза. Когда какой-либо из этих проферментов специфическим образом расщепляется, он становится активным протеолитическим ферментом и расщепляет следующий профермент, и т. д. Поскольку многие из активированных компонентов прочно связываются с мембранами, большинство этих событий происходит на поверхностях клеток. Центральный компонент этого протеолитического каскада — С3. Его активация путем расщепления представляет собой главную реакцию всей цепи активации комплемента. С3 может быть активирован двумя основными путями — классическим и альтернативным. В обоих случаях С3
расщепляется ферментным комплексом, называемым С3-конвертазой. Два разных пути приводят к образованию разных С3-конвертаз, однако обе они образуются в результате спонтанного объединения двух компонентов комплемента, активированных ранее в цепи протеолитического каскада. С3-конвертаза расщепляет С3 на два фрагмента, больший из которых (С3b) связывается с мембраной клетки-мишени рядом с С3-конвертазой; в результате образуется ферментный комплекс еще больших размеров с измененной специфичностью — С5-конвертаза. Затем С5-конвертаза расщепляет С5 и тем самым инициирует спонтанную сборку литического комплекса из поздних компонентов — от С5 до С9. Поскольку каждый активированный фермент расщепляет много молекул следующего профермента, каскад активации ранних компонентов действует как усилитель: каждая молекула, активированная в начале всей цепи, приводит к образованию множества литических комплексов.