ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 09.11.2023
Просмотров: 166
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
Эндополигалактуроназа — фермент, который гидролизует α-1,4-связи в молекуле растворимого пектина (метоксилированной полигалактуроновой кислоты), неупорядоченным, произвольным образом. С возрастанием степени этерификации полигалактуроновой кислоты степень и скорость гидролиза падают, т. к. для проявления каталитической активности фермента требуются свободные карбоксильные группы. Большинство изученных эндополигалактуроназ микроскопических грибов имеют молекулярную массу от 30 000 до 40 000 Да. Оптимальные значения рН колеблются в диапозоне 3,8 — 5,5.
В гидролизе этого типа связи принимает участие и другой фермент — экзополигалактуроназа, который последовательно отщепляет молекулу галактуроновой кислоты, начиная с нередуцирующего конца. Эндополигалактуроназа синтезируется как грибами, так и некоторыми видами бактерий. Они отличаются по своей специфичности к пектинам из различных источников, конечными продуктами реакции, оптимуму рН и другим свойствам.
Для промышленного производства ферментных препаратов пек-толитических ферментов, которые являются комплексными, в качестве продуцентов используют в основном микроскопические (плесневые) грибы, в частности, грибы рода Aspergillus: A. niger, A. wentii, A. oryzae. Бактериальные ферменты в промышленных масштабах не производятся.
Растительные полигалактуроназы, по-видимому, похожи на грибные прлигалактуроназы. Они обнаружены в широком спектре плодов и овощей: помидорах, авокадо, редисе, огурцах, яблоках, грушах, цитрусовых и др. Все они проявляют активность при естественных рН плодов.
313
Применение препаратов пектолитических ферментов в промышленности достаточно обширно. Они используются при производстве фруктовых соковых концентратов и экстрактов, при осветлении соков и вин, при производстве фруктовых и овощных пюре и нектаров.
Целлюлолитические ферменты. Ферментативное разрушение целлюлозы и родственных ей полисахаридов (гемицеллюлозы, лигнина) — сложный процесс, требующий участия комплекса ферментов. Продуцентами такого комплекса целлюлолитических ферментов являются грибы рода Trichoderma, Phanerochaete (Sporotrichum) и Fusarium, a также бактерии рода Clostridium, Cellulomonas и некоторые другие.
Три основных типа целлюлолитических ферментов, продуцируемых микроскопическими грибами, образуют комплекс ферментов, способных осуществить полный гидролиз целлюлозы:
— Эндо-1,4-β-глюконаза или целлюлаза (Н.Ф.3.2.1.4) беспорядочно гидролизует β- 1,4-гликозидные связи. Она не расщепляет целлобиозу, но гидролизует целлодекстрины и производные целлюлозы с высокой степенью замещения, т. к. специфичность этого фермента не высока.
— Экзо-1,4-β-глюконаза или целлобиогидролаза (Н.Ф.3.2.1.91) действует на целлюлозу, отщепляя целлобиозные звенья с нередуцирующего конца цепи. Этот фермент не действует на замещенные производные целлюлозы, что указывает на более высокую субстратную специфичность, чем у эндоглюконазы. Целлобиогидролаза гидролизует целлодекстрины, но не действует на целлобиозу.
— β-Глюкозидаза (Н.Ф.3.2.1.21) расщепляет целлобиозу и целлоолигосахариды до глюкозы. Фермент не действует на целлюлозу и высшие олигосахариды.
Целлюлазная система бактерий существенно проще, чем у грибов, т. к. бактерии образуют только эндоглюконазу и β-глюкозидазу.
Все ферменты целлюлолитического комплекса достаточно хорошо изучены: практически все они являются гликопротеидами, определены их молекулярные массы, изоэлектрические точки, для многих показано наличие множественных форм.
Доказано, что различные ферменты, гидролизующие высокоупорядоченную целлюлозу, действуют в синергизме (рис. 8.10).
Эндоглюконаза атакует аморфные участки целлюлозных фибрилл. В результате образуются новые целлюлозные цепочки, на которые действует целлобиогидролаза, отщепляя целлобиозные звенья с нередуцирующего конца. Синергизм между двумя этими ферментами проявляется в том, что в результате действия эндоглюконазы появляется новый субстрат (более короткие цепочки), на который действует уже целлобиогидролаза. β-Глюкозидаза усиливает гидролиз, расщепляя целлобиозу — конечный продукт и ингибитор этих ферментов.
314
 Рис. 8.10. Схема ферментативного гидролиза целлюлозы
Рис. 8.10. Схема ферментативного гидролиза целлюлозыПрименение целлюлолитических ферментов представляет большой интерес, т. к. ферментативный гидролиз целлюлозосодержащих материалов (древесина, торф, сельскохозяйственные и городские отходы) может обеспечить получение различных биотехнологических продуктов (глюкозы, этанола, ацетона, микробной биомассы). В настоящее время исследования ферментативного расщепления целлюлозы достигли такого уровня, когда имеются все необходимые условия для разработки промышленного процесса и лишь по экономическим причинам он не реализуется на практике.
315
Протеолитические ферменты (Н.Ф.3.4). Основной реакцией, катализируемой протеолитическими ферментами, является гидролиз пептидной связи в молекулах белков и пептидов.
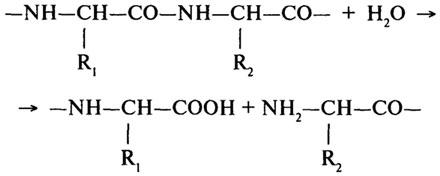
Согласно первоначальной классификации протеолитических ферментов их делили на две группы: протеиназы и пептидазы. При этом считали, что протеиназы действуют на белки, расщепляя их до полипептидов; полипептиды же гидролизуются пептидазами до аминокислот. Более поздняя классификация, до сих пор сохранившая свое значение, основана на схеме, предложенной М. Бергманом и Д. Фрутоном (1937). Согласно этой схеме протеазы разделяются на эндо- и экзопептидазы. Ферменты первой группы (эндопептидазы) могут гидролизовать глубинные пептидные связи и расщеплять молекулу белка на более мелкие фрагменты; ферменты второй группы (экзопептидазы) не могут гидролизовать пептидные связи, находящиеся в середине цепи, и действуют либо с карбоксильного, либо с аминного конца цепи, отщепляя последовательно одну задругой концевые аминокислоты. В связи с этим экзопептидазы подразделяют на аминопептидазы, карбоксипептидазы и дипептидазы.
Аминопептидазы (Н.Ф.3.4.11) катализируют отщепление N-концевых аминокислот.
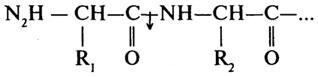
Карбоксипептидазы (Н. Ф. 3.4.12) катализируют отщепление С-концевых аминокислот.
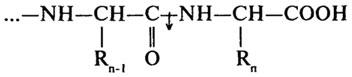
Дипептидазы (Н.Ф.3.4.13) проявляют специфичность к дипептидным субстратам.
Вместе с тем эндопептидазы (протеиназы) разделены на подгруппы, начиная с подподкласса 3.4.21, в первую очередь на основании
316
каталитического механизма (строения активного центра); особенности специфичности используются при этом только для идентификации индивидуальных ферментов в пределах подподкласса:
— сериновые протеиназы (Н.Ф.3.4.21), в активном центре которых функционирует остаток серина и гистидина;
— тиоловые (цистеиновые) протеиназы (Н.Ф.3.4.22), содержат в активном центре SH-группу остатка цистеина;
— кислые (карбоксильные) протеиназы (Н.Ф.3.4.23), в активном центре содержат СООН-группу остатка аспарагиновой кислоты;
— металлопротеиназы (Н.Ф.3.4.24), содержат в активном центре металл, необходимый для проявления их каталитической активности.
Группа протеиназ, о механизме действия которых ничего неизвестно, отнесена к подподклассу 3.4.99.
Субстратная специфичность протеиназ будет определяться следующими факторами: природой аминокислоты, образующей гидролизуемую пептидную связь; радикалами аминокислот, удаленными от гидролизуемой пептидной связи; общей пространственной конформацией молекулы белка, делающей определенные пептидные связи доступными для действия фермента.
Исследование протеолитических ферментов ведется весьма интенсивно. Это происходит вследствие того, что они являются чрезвычайно удобным объектом для изучения структуры белков, активных центров ферментов, механизмов регуляции ферментативной активности и других важных вопросов энзимологии; кроме того, протеазы широко применяются в различных отраслях промышленности: пищевой, сельском хозяйстве, медицине.
Промышленное применение препаратов протеаз будет описано в разделе 8.3. Здесь же мы остановимся на рассмотрении отдельных свойств протеаз, имеющих технологическое значение, предварительно разделив их по типу происхождения на растительные, животные и микробные.
Растительные протеазы. Споры о приоритетности тех или иных ферментных препаратов, полученных из названных источников, ведутся уже достаточно долго. Однако, в связи с ограниченностью животного сырья (внутренние органы крупного рогатого скота и свиней), а также возможной токсичностью микробных препаратов (все они требуют высокой степени очистки, хотя и это не может служить абсолютной гарантией их безопасности), большой интерес представляют растительные протеазы, лишенные указанных недостатков.
Папаин (Н.Ф.3.4.22.2) и химопапаин (Н.Ф.3.4.22.6). Папаин является наиболее применяемым в производстве протеолитическим ферментом. Ферменты папаин и химопапаин — истинные ферменты латекса плодов дынного дерева (Carica papaya).
317
Эти ферменты относятся к группе тиоловых протеиназ, характерной особенностью которых является то, что они активируются сульфгидрильными соединениями — восстановленным глутатионом, цистеином.
Папаин и химопапаин получены в кристаллическом состоянии; их молекулярная масса 20 700 и 36 000 Да, соответственно, а изоэлектрические точки равны 8,75 для папаина и 10,1 для химопапаина.
Оптимальная зона рН для действия папаина зависит от природы гидролизуемого белка и может быть слабокислой, нейтральной или слабощелочной.
Химопапаин имеет большое сходство с папаином, однако можно выделить и некоторые интересные отличия. Например, активность химопапаина при гидролизе гемоглобина и казеина в два раза ниже, чем у папаина; он более термоустойчив, чем папаин, и проявляет хорошую стабильность в кислой среде (рН 2,0).
Папаин обладает довольно широкой специфичностью. Он преимущественно гидролизует вторую пептидную связь, лежащую за карбоксильной группой фенилаланина.
Различные фирмы, как зарубежные, так и отечественные, выпускают ферментные препараты на основе папаина с различной степенью очистки. Возможности их использования обширны: кожевенная промышленность (при обезволашивании и мягчении шкур); кинопроизводство (для растворения желатинового слоя на пленках при их регенерации); парфюмерия (при создании добавок в кремы, лосьоны, зубные пасты); производство синтетических моющих средств (для удаления загрязнений белковой природы); медицина (при лечении воспалительных процессов, ожогов, тромбозов и др.); пищевая промышленность (виноделие, пивоварение, производство спирта, хлебопечение, сыроделие и др.).
Фицин (Н. Ф. 3.4.22.3) и бромелаин (Н. Ф. 3.4.22.5). Фицин выделяют из млечного сока фикусовых растений, например, инжира (Ficus carica). Так же, как и папаин, он относится к тиоловым протеиназам. Еще один тиоловый фермент — бромелаин — получают из свежего сока ананаса (Bromeliacea).
Оба эти фермента имеют сходство с папаином, проявляют наибольшую активность в нейтральной зоне рН, обладают широкой специфичностью, предпочтительно расщепляют пептидные связи, образованные положительно заряженными аминокислотами.
Использование бромелаина и фицина аналогично использованию папаина; в последние годы их производство возросло, и в настоящее время эти ферменты применяют для удаления белковой мути в пиве и для мягчения мяса.
Протеолитические ферменты семян растений. В семенах злаковых и бобовых культур содержится целый комплекс
318
протеолитических ферментов, участвующих в расщеплении запасных белков до аминокислот в процессе прорастания семян. В покоящихся семенах состояние белкового комплекса характеризуется высокой стабильностью и автолиз в водных суспензиях выражен слабо. Поэтому комплекс протеолитических ферментов семян оставался долгое время слабо изученным. Это было связано с методическими трудностями их выделения и очистки. В настоящее время известно, что протеолиз белков в семенах растений осуществляется комплексом ферментов, различающихся по своим функциям, механизму действия и другим показателям. Некоторые из этих ферментов были выделены в виде высокоочищенных препаратов и подробно охарактеризованы. Так, например, из семян пшеницы были выделены несколько типов протеолитических ферментов, различающихся по оптимуму рН: кислые протеиназы с оптимумом рН 3,7 — 4,0; нейтральные протеиназы с оптимумом рН 6,5 — 7,0; щелочные протеиназы с оптимумом рН > 8,0.
Из трех групп протеиназ наибольшего внимания технологов заслуживают нейтральные протеиназы. По активности они в несколько раз превосходят кислые и в условиях теста способны эффективно расщеплять белки клейковины. Одна из особенностей нейтральных протеиназ состоит в том, что они не растворяются в водных, солевых и буферных растворах. Они прочно связаны с белками клейковинного комплекса и извлекаются при частичном растворении клейковины в щелочном растворе. Максимальное извлечение нейтральных протеиназ происходит при обработке измельченного зерна, муки или лиофилизированной клейковины 0,35%-м раствором карбоната натрия. При подкислении щелочного раствора нейтральные протеиназы выпадают в осадок, а в надосадочной жидкости остаются их белковые ингибиторы. Таким образом, в созревших семенах пшеницы нейтральные протеиназы и их белковые ингибиторы образуют единый неактивный комплекс, связанный с клейковиной. Соотношение активности протеиназ и их ингибиторов в созревшем зерне определяет стабильность белкового комплекса, его устойчивость в процессе тестоведения.