Файл: Занятие 1 Тема 1 Растворы и их роль в жизнедеятельности Дайте краткий ответ на теоретические вопросы.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 23.11.2023
Просмотров: 1041
Скачиваний: 2
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
Лизофосфолипиды – продукт гидролиза фосфолипидов фосфолипазой А2, образуются при определенных стимулах, вызывающих в клетке синтез эйкозаноидов (простагландинов, лейкотриенов).Гораздо более редким является кардиолипин – структурный фосфолипид в мембране митохондрий.Плазмалогены при С1 содержат высший спирт вместо жирной кислоты. Они участвуют в построении структуры мембран, составляют до 10% фосфолипидов мозга и мышечной ткани.Строение менее распространенных фосфолипидовСфингофосфолипидыОсновным представителем у человека являются сфингомиелины – основное их количество расположено в сером и белом веществе головного и спинного мозга, в оболочке аксонов периферической нервной системы, есть в печени, почках, эритроцитах и других тканях. В качестве жирных кислот выступают насыщенные и мононенасыщенные.В нервной ткани сфингомиелин участвует в передаче нервного сигнала по аксонам. В последние годы активно разрабатывается роль сфинголипидов в регуляциивнутриклеточных процессов в качестве источника вторичного мессенджера церамида.Решите задачи:Задача №1В двух пробирках содержатся пробы подсолнечного и сливочного масла одинаковой массы. В лаборатории имеются бром, этиловый спирт, четыреххлористый углерод, едкий натр. Требуется распознать содержимое пробирок.С помощью бромной воды в подсолнечном масле непредельные кислоты, а в сливочном предельные. В подсолнечном масле раствор брома будет обесцвечиваться. А в сливочном нетВопросы:1. Остатки каких кислот преимущественно входят в состав сливочного и подсолнечного масел?2. При помощи какого реагента можно распознать содержимое пробирок?3. В каком растворителе следует проводить реакцию?4. В чем заключается предлагаемая реакция и каковы ее визуальные признаки?5. Как по результатам анализа определить содержимое пробирок?Задача 2.Приготовление майонезаВ процессе приготовления майонеза фосфатидилхолин (лецитин) из яичных желтков переходит в растительное масло, что стабилизирует соус и не позволяет ему расслаиваться. Объясните, почему это происходит. Ответ поясните.
Ответьте на тесты:1. При промышленном гидрировании растительных масел образуется

Под трансаминированием подразумевают реакции межмолекулярного переноса аминогруппы (NH2—) отаминокислоты на α-кетокислоту без промежуточного образования аммиака. Впервые реакции трансаминиро-вания (прежнее название «переаминирование») были открыты в 1937 г. советскими учеными А.Е. Браунштейном и М.Г. Крицман при изучении дезаминирования глутаминовой кислоты в мышечной ткани. Было замечено, что при добавлении к гомогенату мышц глутаминовой и пировиноградной кислот образуются α-кетоглутаровая кислота иаланин без промежуточного свободного аммиака; добавление аланина и α-кетоглутаровой кислоты приводило к образованию соответственно пировиноградной и глутаминовой кислот.
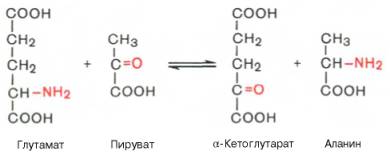
Реакции трансаминирования являются обратимыми и, как выяснилось позже, универсальными для всех живыхорганизмов. Эти реакции протекают при участии специфических ферментов, названных А.Е. Браун-штейном аминоферазами (по современной классификации, аминотранс-феразы, или трансаминазы). Теоретически реакциитрансаминиро-вания возможны между любой амино- и кетокислотой, однако наиболее интенсивно они протекают в том случае, когда один из партнеров представлен дикарбоновой амино- или кетокислотой. В тканях животных и умикроорганизмов доказано существование реакций трансаминирования между монокарбоновыми амино- икетокислотами. Донорами NН2-группы могут также служить не только α-, но и β-, γ- и ω-аминогруппы рядааминокислот. В лаборатории А. Майстера доказано, кроме того, трансами-нирование глутамина и аспарагина скетокислотами в тканях животных.
В переносе аминогруппы активное участие принимает кофермент транс-аминаз пиридоксальфосфат (производноевитамина В6; см. главу 5), который в процессе реакции обратимо превращается в пиридоксаминфосфат.
Механизм реакциитрансаминирования. Общую теорию механизма ферментативного трансаминированияразработали советские ученые А.Е. Браун-штейн и М.М. Шемякин. Одновременно подобный механизм был предложен американскими биохимиками Э. Снеллом и Д. Метцлером. Все трансаминазы (как и декарбоксилазыаминокислот) содержат один и тот же кофермент – пиридоксальфосфат. Для реакций трансаминирования харак -терен общий механизм. Специфичность трансаминаз обеспечивается белковым компонентом. Ферментытрансаминирования катализируют перенос NH
2-группы не на α-кетокислоту, а сначала на кофермент пиридоксаль-фосфат. Образовавшееся промежуточное соединение (шиффово основание) подвергается внутримолекулярным превращениям (лабилизация α-водо-родного атома, перераспределение энергии связи), приводящим к освобождению α-кетокислоты и пиридоксаминфосфата; последний на второй стадии реакции реагирует с любой другой α-кетокислотой, что через те же стадии образования промежуточных соединений (идущих в обратном направлении) приводит к синтезу новой аминокислоты и освобождению пиридоксальфосфата. Опуская промежуточные стадии образования шиффовых оснований, обе стадии реакции трансаминирования можно представить в виде общей схемы:
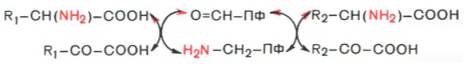
Более подробно механизм действия трансаминаз представлен на рис. 12.3.
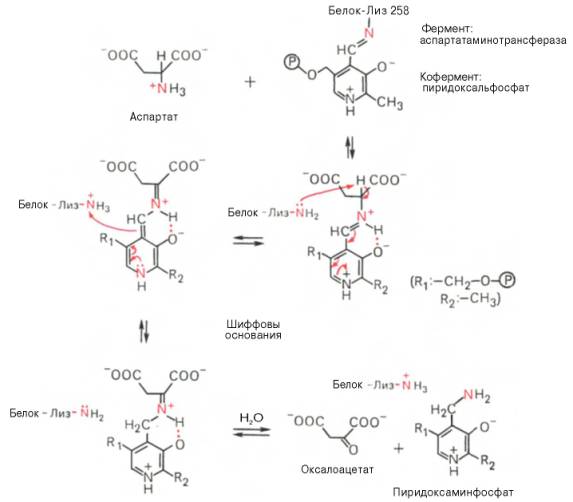
В связи с тем что во всех пиридоксалевых ферментах (включая транс-аминазы) карбонильная группа кофермента(—СНО) оказалась связанной с ε-аминогруппой лизина белковой части, в классический механизм реакциитрансаминирования А.Е. Браунштейн и Э. Снелл внесли следующее дополнение. Оказалось, что взаимодействие между субстратом, т.е. L-амино-кислотой (на рисунке – аспартат), и пиридоксальфосфатом происходит не путемконденсации с выделением молекулы воды, а путем реакции замещения, при которой NH2-группа субстратавытесняет ε-NН2-группу лизина в молекуле ферментного белка, что приводит к формированию пиридоксальфосфатного комплекса.
Решите задачи:Задача №1В лабораторию на анализ поступил пептид неизвестного строения. В лаборатории имеются следующие реактивы: нингидрин; п-диметиламино-бензальдегид; азотная кислота; 2,4-динитрофторбензол. Необходимо идентифицировать N-концевую α-аминокислоту.Для определения природы N-концевой аминокислоты предложен ряд методов, в частности метод Сэнджера (F. Sanger), основанный на реакции арилирования полипептида 2,4-динитрофторбензолом (ДНФБ), что приводит к образованию окрашенного в желтый цвет 2,4-динитрофенильного производного N-концевой аминокислоты . Растворполипептида обрабатывают ДНФБ, который взаимодействует со свободной NH2-группой N-концевой аминокислотыпептида.После кислотного гидролиза продукта реакции – динитрофенилпептида только одна N-концевая аминокислотаоказывается связанной с реактивом в виде 2,4-динитрофениламинокислоты (стабильной при гидролизе). В отличие от других образовавшихся при гидролизе полипептида свободных аминокислот она желтого цвета. Ее идентифицируют методом хроматографии.Для определения N-концевой аминокислоты значительно более широко применяется фенилтиогидантоиновый метод Эдмана благодаря своей высокой чувствительности и возможности многократного использования в одной и той же пробе. Фенилизотиоцианат реагирует со свободной α-NH2-группой N-концевой аминокислоты полипептида с образованием фенилтиокарбамоилпептида.Обработка продукта реакции кислотой приводит к циклизации и освобождению фенилтиогидантоина N-концевойаминокислоты, природу которого устанавливают хроматографически. Укороченный на одну аминокислотуполипептид подвергают дальнейшему анализу.Эту процедуру ступенчатого расщепления пептида с N-конца можно повторять многократно, идентифицируя последовательно одну аминокислоту за другой. Метод Эдмана используется в качестве химической основы для определения первичной структуры белков и пептидов. Он реализован в специальном приборе – секвенаторе (от англ. sequence – последовательность), работающем в автоматическом режиме и позволяющем определить последовательность аминокислот с N-конца пептида до 50–60 аминокислотных остатков.
Ответьте на тесты:1. При промышленном гидрировании растительных масел образуется
-
синтетические масла -
синтетическое топленое масло -
твердый жир -
синтетическая сметана -
синтетический белок
-
гидролиза жира -
гидрогенизации жира -
гидрогенизации растительного масла в молоке -
термической обработки молока -
гидролиза растительного масла в молоке
-
альдегиды -
карбоновые кислоты -
гликоли -
+альдегиды и карбоновые кислоты -
перекиси
-
триацилглицерины -
воска -
фосфолипиды, сфинголипиды, гликолипиды -
только фосфолипиды -
только сфинголипиды
-
Понятие о трансаминировании, строение кофактора ПАЛФ (роль витамина В6 в этом процессе). Механизм трансаминирования. Схема реакции на примере АЛТ (аланинаминотрансферазы). Биологическая роль трансаминирования.
- 1 2 3 4 5 6
Трансаминирование аминокислот
Под трансаминированием подразумевают реакции межмолекулярного переноса аминогруппы (NH2—) отаминокислоты на α-кетокислоту без промежуточного образования аммиака. Впервые реакции трансаминиро-вания (прежнее название «переаминирование») были открыты в 1937 г. советскими учеными А.Е. Браунштейном и М.Г. Крицман при изучении дезаминирования глутаминовой кислоты в мышечной ткани. Было замечено, что при добавлении к гомогенату мышц глутаминовой и пировиноградной кислот образуются α-кетоглутаровая кислота иаланин без промежуточного свободного аммиака; добавление аланина и α-кетоглутаровой кислоты приводило к образованию соответственно пировиноградной и глутаминовой кислот.
Реакции трансаминирования являются обратимыми и, как выяснилось позже, универсальными для всех живыхорганизмов. Эти реакции протекают при участии специфических ферментов, названных А.Е. Браун-штейном аминоферазами (по современной классификации, аминотранс-феразы, или трансаминазы). Теоретически реакциитрансаминиро-вания возможны между любой амино- и кетокислотой, однако наиболее интенсивно они протекают в том случае, когда один из партнеров представлен дикарбоновой амино- или кетокислотой. В тканях животных и умикроорганизмов доказано существование реакций трансаминирования между монокарбоновыми амино- икетокислотами. Донорами NН2-группы могут также служить не только α-, но и β-, γ- и ω-аминогруппы рядааминокислот. В лаборатории А. Майстера доказано, кроме того, трансами-нирование глутамина и аспарагина скетокислотами в тканях животных.
В переносе аминогруппы активное участие принимает кофермент транс-аминаз пиридоксальфосфат (производноевитамина В6; см. главу 5), который в процессе реакции обратимо превращается в пиридоксаминфосфат.
Механизм реакциитрансаминирования. Общую теорию механизма ферментативного трансаминированияразработали советские ученые А.Е. Браун-штейн и М.М. Шемякин. Одновременно подобный механизм был предложен американскими биохимиками Э. Снеллом и Д. Метцлером. Все трансаминазы (как и декарбоксилазыаминокислот) содержат один и тот же кофермент – пиридоксальфосфат. Для реакций трансаминирования харак -терен общий механизм. Специфичность трансаминаз обеспечивается белковым компонентом. Ферментытрансаминирования катализируют перенос NH
2-группы не на α-кетокислоту, а сначала на кофермент пиридоксаль-фосфат. Образовавшееся промежуточное соединение (шиффово основание) подвергается внутримолекулярным превращениям (лабилизация α-водо-родного атома, перераспределение энергии связи), приводящим к освобождению α-кетокислоты и пиридоксаминфосфата; последний на второй стадии реакции реагирует с любой другой α-кетокислотой, что через те же стадии образования промежуточных соединений (идущих в обратном направлении) приводит к синтезу новой аминокислоты и освобождению пиридоксальфосфата. Опуская промежуточные стадии образования шиффовых оснований, обе стадии реакции трансаминирования можно представить в виде общей схемы:
Более подробно механизм действия трансаминаз представлен на рис. 12.3.
В связи с тем что во всех пиридоксалевых ферментах (включая транс-аминазы) карбонильная группа кофермента(—СНО) оказалась связанной с ε-аминогруппой лизина белковой части, в классический механизм реакциитрансаминирования А.Е. Браунштейн и Э. Снелл внесли следующее дополнение. Оказалось, что взаимодействие между субстратом, т.е. L-амино-кислотой (на рисунке – аспартат), и пиридоксальфосфатом происходит не путемконденсации с выделением молекулы воды, а путем реакции замещения, при которой NH2-группа субстратавытесняет ε-NН2-группу лизина в молекуле ферментного белка, что приводит к формированию пиридоксальфосфатного комплекса.
-
Понятие о дезаминировании АМК. Виды дезаминирования: окислительное (глутаминовая кислота), гидролитическое (серин), внутримолекулярное (гистидин), восстановительное (аланин). Биологическая роль дезаминирования.
-
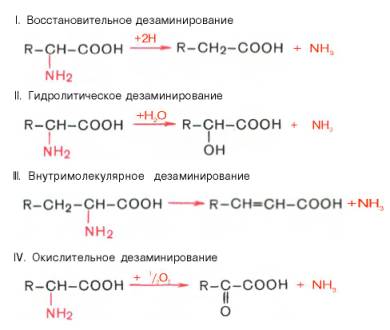
Доказано существование 4 типов дезаминирования аминокислот (отщепление аминогруппы). Выделены соответствующие ферментные системы, катализирующие эти реакции, и идентифицированы продукты реакции. Во всех случаях NH2-группа аминокислоты освобождается в виде аммиака. -
-
Помимо аммиака, продуктами дезаминирования являются жирные кислоты, оксикислоты и кетокислоты. Для животных тканей, растений и большинства аэробных микроорганизмов преобладающим типом реакций является окислительное дезаминирование аминокислот, за исключением гис-тидина, подвергающегося внутримолекулярномудезаминированию.
Решите задачи:Задача №1В лабораторию на анализ поступил пептид неизвестного строения. В лаборатории имеются следующие реактивы: нингидрин; п-диметиламино-бензальдегид; азотная кислота; 2,4-динитрофторбензол. Необходимо идентифицировать N-концевую α-аминокислоту.Для определения природы N-концевой аминокислоты предложен ряд методов, в частности метод Сэнджера (F. Sanger), основанный на реакции арилирования полипептида 2,4-динитрофторбензолом (ДНФБ), что приводит к образованию окрашенного в желтый цвет 2,4-динитрофенильного производного N-концевой аминокислоты . Растворполипептида обрабатывают ДНФБ, который взаимодействует со свободной NH2-группой N-концевой аминокислотыпептида.После кислотного гидролиза продукта реакции – динитрофенилпептида только одна N-концевая аминокислотаоказывается связанной с реактивом в виде 2,4-динитрофениламинокислоты (стабильной при гидролизе). В отличие от других образовавшихся при гидролизе полипептида свободных аминокислот она желтого цвета. Ее идентифицируют методом хроматографии.Для определения N-концевой аминокислоты значительно более широко применяется фенилтиогидантоиновый метод Эдмана благодаря своей высокой чувствительности и возможности многократного использования в одной и той же пробе. Фенилизотиоцианат реагирует со свободной α-NH2-группой N-концевой аминокислоты полипептида с образованием фенилтиокарбамоилпептида.Обработка продукта реакции кислотой приводит к циклизации и освобождению фенилтиогидантоина N-концевойаминокислоты, природу которого устанавливают хроматографически. Укороченный на одну аминокислотуполипептид подвергают дальнейшему анализу.Эту процедуру ступенчатого расщепления пептида с N-конца можно повторять многократно, идентифицируя последовательно одну аминокислоту за другой. Метод Эдмана используется в качестве химической основы для определения первичной структуры белков и пептидов. Он реализован в специальном приборе – секвенаторе (от англ. sequence – последовательность), работающем в автоматическом режиме и позволяющем определить последовательность аминокислот с N-конца пептида до 50–60 аминокислотных остатков.