Файл: 1. Анатомия сосудов головного мозга. Виллизиев круг. Кровоснабжение и отток. Бассейны. Анатомия кровоснабжения головного мозга.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 30.11.2023
Просмотров: 148
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
-
Вертебробазилярный бассейн. Топографическое расположение. Важность бассейна. Близость к сосудам и нервам.
Вертебро-базилярный бассейн формируется двумя позвоночными артериями, берущими начало от подключичных артерий и их ветвями (базилярной, задними мозговыми и мозжечковыми артериями. Артерии вертебробазилярного бассейна осуществляют кровоснабжения около 15-25% головного мозга.
Позвоночные артерии (ПА) на шее проходят через костный канал, образованный отверстиями поперечных отростков VI—II шейных позвонков. Выйдя из отверстия поперечного отростка позвонка СII, ПА делает изгиб, поворачивая кнаружи, а затем вверх, проходит сквозь отверстие поперечного отростка атланта и через большое затылочное отверстие проникает в полость черепа. Субокципитальная часть ПА образует извитость, которая при поворотах и сгибаниях головы предотвращает ее резкое растяжение. Прободая атлантозатылочную мембрану (membranaatlanto-occipitalis) и твердую мозговую оболочку, обе ПА, сближаясь, подходят к нижней поверхности продолговатого мозга. На его границе с мостом они соединяются, образуя базилярную артерию (a. basilaris). От ПА уже в полости черепа отходят ветви, которые, соединяясь, образуют переднюю спинномозговую артерию (a. spinalisanterior). Кроме того, от ПА до места их слияния отходят менингеальные ветви и нижняя задняя мозговая артерия (a. cerebelliinferiorposterior), участвующие в снабжении кровью продолговатого мозга и мозжечка.
Базилярная артерия на границе моста со средним мозгом делится на две задние мозговые артерии (ЗМА). От базилярной артерии и задних мозговых артерий отходят: передняя нижняя мозжечковая артерия (a. cerebelliinferioranterior) и отходящая от нее ветвь внутреннего слухового прохода (a. labyrinthi), верхняя мозжечковая артерия (a. cerebellianterior).
Этот бассейн является общим для обеих позвоночных артерий и поэтому нарушение кровообращения только по одной позвоночной артерии может вызвать симптомы из всей территории вертебробазилярного бассейна мозга. Слуховые центры и нервы, вестибулярные ядра, нижние части мозжечка кровоснабжаются позвоночной артерией той же самой стороны еще до слияния их в основную артерию. Потому звон или шум в ушах, понижение или потеря слуха характерны для уха на той же самой стороне, как и поврежденная позвоночная артерия.
-
ЗМА (задняя мозговая артерия), ветви. Топографическое расположение. Важность бассейна. Близость к другим сосудам и нервам.
Задняя мозговая артерия (a. cerebriposteror) огибает ножку мозга и идет к наружному коленчатому телу. Далее направляется через отверстие намета мозжечка к височной доле, проходит по внутреннему краю ее нижней поверхности и достигает полюса затылочной доли.
Сосуд условно подразделяется на три части, которые еще называют сегментами задней мозговой артерии.
-
предкоммуникационная часть; -
посткоммуникационная часть; -
конечная, или корковая, часть.
Предкомуникационная часть (P1 сегмент задней мозговой артерии) - это участок сосуда, расположенный до места впадения в него задней соединительной артерии. От нее отходят такие ветви, как медиальная задняя хориоидальная, парамедианная мезэнцефалическая, а также задняя таламоперфорирующая артерии. Они несут кровь ядрам таламуса и коленчатому телу (медиальной его части).
Посткоммуникационная часть (P2 сегмент) - это участок, размещенный после впадения в сосуд задней соединительной артерии. От него отходят следующие ветви задней мозговой артерии: таламогеникулятная, педункулярные перфорантные и латеральная задняя хориоидальная артерии. Они также снабжают кровью коленчатое тело, но срединную его часть. Кроме того, эти сосуды несут кислород и питательные вещества к части среднего мозга, ядрам и подушке таламуса, боковой стенке первого и второго желудочков.
Конечная часть (P3 и P4 сегменты) несет кровь к коре головного мозга. Она дает переднюю и заднюю височные, шпорную и теменно-височные ветви. В большинстве случаев кровь от конечного сегмента поступает к отделам коры до сильвиевой борозды.
Основные ветви задней мозговой артерии — заднемедиальные центральные артерии, таламические ветви, медиальные задние ворсинчатые ветви, ножковые ветви, латеральная и медиальная затылочные артерии, шпорная ветвь, направляющаяся к корковому полю 17, по Бродману (к корковому концу зрительного анализатора). Небольшая часть этого поля получает кровь из СМА, чем и объясняется сохранность макулярного зрения при гомонимной гемианопсии, обусловленной окклюзией ЗМА.
В месте прохождения ЗМА через тенториальное отверстие может сдавливаться при вклинении височной доли.
В глубину полушария уходят 3 ветви ЗМА: таламоперфорирующая, премаммилярная и таламоколенчатая. Они снабжают кровью задненижнюю часть зрительного бугра и гипоталамической области, утолщение мозолистого тела, зрительный венец и льюисово тело. Самая крупная из этих ветвей — задняя ворсинчатая артерия, которая кончается в сосудистом сплетении III мозгового желудочка.
Важность в том, что клинические проявления нарушения тока крови в ЗМА зависят от локализации повреждения. Это могут быть симптомы поражения среднего мозга, таламуса, затылочной и теменной области коры головного мозга.
7. Анатомия и физиология мозгового кровообращения. Особенности мозгового кровотока у здорового человека. Энергетический метаболизм головного мозга.
8. Механизмы регуляции (общие и местные) и саморегуляции. Феномен Остроумова-Бейлиса. Феномен «обкрадывания» и обратного «обкрадывания». Клиническое значение.
Физиология мозгового кровообращения.
Важной особенностью мозгового кровообращения является его определенная автономность, регуляция которого осуществляется через сложную систему, в которую входят следующие компоненты:
-
Мышечная системы сосудистой стенки (эффект Остроумова-Бейлисса); -
Состояние метаболизма мозговой ткани; -
Напряжение кислорода (O2); -
Напряжение углекислого газа (СО2); -
Состояние кислотно-основного состояния и рН ткани мозга, количество молочной и пировиноградной кислоты; -
Нервная регуляция; -
Анатомические особенности сосудов головного мозга.
Осуществление регуляции мозгового кровотока за счет колебаний внутрисосудистого давления происходит за счет его прямого влияния на тонус гладких мышц мозговых артерий. Реакция мышечных клеток па изменения внутрисосудистого давления, впервые описанная А. Остроумовым (1876), а затем детально изученная Bayliss (1902, 1923), выражается в вазоконстрикции при повышении артериального давления и вазодилатации в случаях его падения.
Механизм этой реакции считается миогенным, так как сокращение гладких мышц сосудов возникает при растяжении их стенки, даже если для этого берется изолированный сегмент артерии. Таким образом, при повышении уровня общего артериального давления происходит сужение сосудов мозга, при снижении — их расширение.
Феномен саморегуляции мозгового кровотока осуществляется в определенных границах при колебаниях общего артериального давления между 60 и 180 мм рт. ст. (табл. 1).
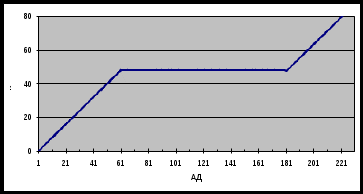
Табл.1: Схема взаимоотношений между мозговым кровотоком и артериальным давлением (по Lassen, 1959). МК – мозговой кровоток в миллилитрах на 100 г в 1 мин; АД – артериальное давление в миллиметрах ртутного столба.
Саморегуляторный ответ сосудов мозга, в основе которого лежит эффект Остроумова — Бейлисса, длится очень короткое время, не более чем 30 с. Затем начинает проявляться влияние сдвигов метаболизма ткани мозга.
В последние годы в связи с исследованием влияния внутричерепного давления на мозговой кровоток и его саморегуляцию специальному обсуждению подвергаются представления о так называемом перфузионном мозговом давлении. Под перфузионным мозговым давлением понимается разность между средним артериальным давлением и внутричерепным давлением. Для сохранения саморегуляции мозгового кровотока и отсутствия его редукции при повышении внутричерепного давления необходимо, чтобы перфузионное мозговое давление не снижалось ниже 40 мм. рт. ст.
Определенное влияние на мозговой гемодинамики оказывают метаболические сдвиги ткани мозга. При снижении содержания в артериальной крови кислорода ниже 60-50% от исходной величины возникает расширение мозговых сосудов. В ответ на гипероксию мозговые сосуды суживаются и мозговой кровоток заметно снижается.
Углекислота оказывает обратное влияние на тонус сосудов мозга и мозговой кровоток. Уменьшение содержания углекислоты в артериальной крови и ткани мозга вызывает сосудосуживающий эффект и снижение мозгового кровотока. Гиперкапния и накопление углекислоты в ткани мозга являются одним из самых сильных сосудорасширяющих факторов, увеличивающих мозговой кровоток. Влияние разных концентраций углекислоты во вдыхаемом воздухе и в артериальной крови на тонус сосудов мозга и мозговой кровоток весьма вариабельно. Например, при ингаляции 5% смеси углекислого газа мозговой кровоток может увеличиваться на 50% .
Влияние кислотно-основного состояния можно рассмотреть на примере данных работ М. Е. Маршака (1965) и других, при активизации нервных клеток, вызванной пробуждением, освещением в темной комнате, при стрихнинизации коры мозга увеличивается потребление кислорода и глюкозы, в связи с чем падает рО
2, возрастает продукция рСО2 и лактата, снижается количество АТФ и креатинфосфата, а количество неорганического фосфата увеличивается. Вслед за указанными сдвигами в исследуемой части мозга падает тонус артерий, они расширяются, и мозговой кровоток возрастает. В результате компенсируется недостаток кислорода и удаляется избыточное количество углекислоты и других продуктов метаболизма.
Обратные явления наблюдаются при наркозе, когда угнетается дыхание ткани мозга, снижается количество образующейся молочной кислоты и углекислоты, повышается содержание АТФ и креатинфосфата, а количество неорганического фосфата снижается. После нормализации метаболизма мозговой кровоток начинает возвращаться к исходному уровню.
Велика роль в регуляции мозгового кровообращения принадлежит рефлексогенным влияниям артериальной синокаротидной зоны и возбуждению симпатической нервной системы, которые вызывают рефлекторный спазм крупных и средних артериальных стволов как на поверхности, так и внутри мозгового вещества.
Анатомические особенности представлены в виде физиологических приспособлений, регулирующих величину и скорость пульсовых артериальных волн. Удар пульсовой волны амортизируется благодаря изгибам крупных (внутренних сонных, позвоночных) артерий, сложному строению артериальной сети, наличию анастомозов, особенностям хода артерий, снабжающих кровью подкорковые узлы, а также отхождению от артерий мягкой мозговой оболочки ветвей, идущих радиально в мозг и питающих его поверхностные структуры, в том числе кору и прилежащее к ней белое вещество.
Повреждения и отек мозга, нарушение кровоснабжения вследствие патологии сосудистой системы ограничивают возможности саморегуляции мозгового кровотока следующим образом:
-
Сосудорасширяющие влияния приводят к увеличению притока крови только в здоровые области мозга, тем самым анемизируя патологический очаг – синдром внутримозгового обкрадывания; -
Сосудосуживающие влияния, уменьшая кровоснабжение здоровых тканей, перераспределяют кровоток в пользу очага повреждения – извращенный синдром внутримозгового «обкрадывания» (синдром Робин-Гуда); -
При прекращении кровотока в проксимальных отделах дуги аорты дистальнее отхождения левой общей сонной артерии, но проксимальнее отхождения левой позвоночной артерии может возникать «подключичный феномен обкрадывания», когда левая позвоночная артерия играет роль коллатерали для левой руки -
Энергетический метаболизм головного мозга. -
Мозговая ткань беспрерывно активно функционирует, а ее потребность в кислороде больше, чем у других тканей и органов. Масса мозга составляет приблизительно 2% массы тела — 1350—1450 г. За минуту через мозг проходит около 800 мл крови, что составляет 15—20% сердечного выброса, при этом в мозге нет запасов кислорода и единственного субстрата, необходимого для выработки энергии, — глюкозы. Потому необходимо постоянное поступление глюкозы в количестве 75—100 мг/мин или 125 г/сут. В метаболических процессах в мозговой ткани, направленных на выработку энергии, преобладающим (90%) является аэробный гликолиз, в цикле которого образуется 36 молекул АТФ. Тогда как за счет анаэробного гликолиза обеспечивается 10% энергетических процессов, синтезирующего только 2 молекулы АТФ. Постоянное обеспечение мозга АТФ требуется для поддержания целостности и функциональной активности его ткани, в частности сохранения необходимого соотношения главных внутриклеточных катионов — ионов калия (К+) и основных внеклеточных катионов — ионов натрия (Na+) и кальция (Са2+). -
Кровь, поступающая в мозг и содержащая нужное количество глюкозы, для обеспечения аэробных метаболических процессов должна быть и достаточно оксигенированной. Общий мозговой кровоток у здорового человека составляет 50—55 мл на 100 г мозговой ткани в 1 мин (от 45 до 75 мл). В таком случае в мозг попадает около 20% поступающего в организм кислорода. Общее потребление мозгом кислорода, т.е. скорость церебрального метаболизма кислорода, составляет 3,3—3,5мл/100 г/мин. Таким образом, головной мозг в минуту потребляет около 45 мл кислорода. -
Для обеспечения достаточного кровотока в мозге и полноценного снабжения его необходимыми ингредиентами должны быть в норме общее кровообращение, состав крови, ее насыщенность кислородом и многие другие факторы которые будут рассмотрены в соответствующих разделах.