ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 11.12.2023
Просмотров: 105
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
реттелмейді. Мәселен, мысалы, стресс кортиколибериннің босатылуын тудырады, сәйкесінше кортизол секрециясын арттыратын реакциялардың барлық сериясы. Сонымен қатар, кортизол секрециясы күнделіктімуаққа бағынады; ол оянғанда өте жоғары, бірақ ұйқы кезінде біртіндеп минималды деңгейге дейін төмендейді. Бақылау механизмдеріне гормонның метаболизм жылдамдығы және олардың белсенділігін жоғалту жатады. Ұқсас реттеу жүйелері басқа гормондарға да әсер етеді.Орталық жүйке жүйесі гормондардың секрециясын реттеуде маңызды. Эндокриндік бездердің қызметін үйлестіретін және бақылайтын ОЖЖ-нің маңызды бағыттарының бірі гипоталамус болып табылады, онда аденогипофиз гормондарының синтезі мен секрециясын реттеуге қатысатын
нейросекреторлық ядролар мен орталықтар локализацияланған. Гипоталамус-гипофизді реттеу кері байланыс принципі бойынша жұмыс істейтін механизмдер арқылы жүзеге асырылады, онда өзара әрекеттесудің әртүрлі деңгейлері айқын ерекшеленеді (сурет. 1).
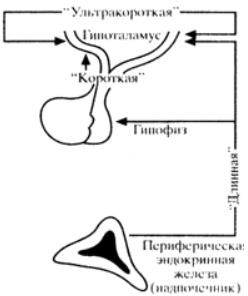
Сурет 2. Керібайланысдеңгейлері.
"Ұзын" керібайланыс тізбегі перифериялық эндокриндік бездің гипофиз және гипоталамус орталықтарымен (ОЖЖ-ніңсупрагипоталамикалық және басқа аймақтарымен)әсер ету " керібайланыс тізбегі гипофиздің тропиктік гормонының жоғарылауы (мысалы, ACTH) гипофизотропты гормонның (бұл жағдайда кортиколиберин) секрециясы мен бөлінуін модуляциялайты және өзгертетін өзара әрекеттесуді білдіреді."Ультра қысқа" керібайланыстізбегі- біргипофизотропты гормонның бөлінуі басқа гипофизотропты гормонның бөлінуі мен бөліну процестеріне әсер ететін гипоталамус ішіндегі өзара әрекеттесутүрі. Кері байланыстың бұл түрікез- келген эндокриндік безде болады. Сонымен, осы нейрондардың аксондары арқылы және жасуша аралық өзара әрекеттесу арқылы
(жасушадан жасушаға) окситоцин немесе вазопрессиннің бөлінуі осы гормондарды шығаратын нейрондардың белсенділігін өзгертеді. Тағы бірмысал, пролактиннің бөлінуі және оның васкулярлық кеңістіктерге диффузиясык өршілес лактотрофтарға әсер етеді, соданкейін пролактин секрециясының тежелуі."Ұзын "және" қысқа "керібайланыс тізбектері" жабық " типті жүйелер ретінде жұмыс істейді, яғни олар өзін-өзі реттейтін жүйелер.Алайда, олар ішкі және
сыртқы сигналдарға жауап береді, қысқа уақытқа өзін- өзі реттеу принципін өзгертеді (мысалы, стресс кезінде және т.б.). Сонымен қатар,бұл жүйелерге күн мен түннің өзгеруіне байланысты биологиялық циркадиялық ритақты қолдайтын механизмдер әсер етеді. Күндізгі-түнгі меақтуралы ақпарат Күн-түн механизмінен басқа, осы "сағаттардың" қызметіне басқа реттеушілер қатысады (Дене температурасының өзгеруі, демалу, ұйқы және т.б.).
2.2. Көмірсу, липид, ақуыз және су - тұз алмасуының гормоналды реттелуі Тірі организмнің негізгі энергетикалық ресурстары — көмірсулар мен
майлар ферменттік катаболикалық түрлендірулер арқылы жасушаларда оңай алынатын потенциалдық энергияның жоғарық орына ие. Көмірсулар мен май алмасуларының, сондай-ақ гликолиз өнімдерінің биологиялық тотығу процесінде бөлінетін Энергия негізінен синтезделген АТФ фосфат байланыстарының химиялық энергиясына айналады.АТФ-да жинақталған макроэргиялық байланыстардың химиялық энергиясы, өз кезегінде, әртүрлі жасушалық жұмыстарға жұмсалады. — Электрохимиялық градиенттерді құружәне қолдау, бұлшық еттердің жиырылуы, секреторлық және кейбір көлік процестері, ақуыз, май қышқылдарының биосинтезі және т. б. "отын" функциясынан басқа, көмірсулар мен майлар ақуыздармен бірге маңызды құрылыс, пластикалық жеткізушілер рөлін атқарады. Жасушаның негізгі құрылымдарына кіретін материалдар-нуклеин қышқылдары, қарапайым ақуыздар, гликопротеидтер, бірқатар липидтер және т.б. Көмірсулар мен майлардың ыдырауына байланысты синтезделген
АТФ жасушаларды жұмыс
істеугеқажетті энергиямен қамтамасыз етіп қана қоймайды, сонымен қатар camp түзілу көзі болып табылады, сонымен қатар көптеген ферменттердің белсенділігін, құрылымдық ақуыздардың күйін реттеуге қатысады, олардың фосфорлануын қамтамасыз етеді.Жасушалар тікелей тастайтын көмірсулар мен липидті субстраттар-моносахаридтер (ең алдымен глюкоза) және эфирленбеген май қышқылдары (HEFA), сондай-ақ кейбір тіндерде кетон денелері. Олардың көздері-көмірсулардың гликоген түрінде және бейтарап майлар түрінде ағзаларға салынған ішектен сіңетін Тамақ өнімдері
липидтер, сондай-ақ көмірсулар түзетін аминқышқылдары мен глицерин (глюконеогенез). Омыртқалылардағы депозиттік органдарға бауыр мен майлы (адипозды) тін, глюконеогенез органдарына — бауыр мен бүйрек жатады. Жәндіктерде май ағзасы депозиттік орган болып табылады. Сонымен қатар, глюкоза мен HEHA көздері жұмыс істейтін жасушада сақталған немесе өндірілген кейбір қосалқы немесе басқа өнімдер болуы мүмкін. Көмірсулар мен май алмасуының әртүрлі жолдары мен кезеңдері көптеген өзара әсерлермен байланысты. Бұл алмасу процестерінің бағыты мен қарқындылығы бірқатар сыртқы және ішкі факторларға байланысты. Оларға, атап айтқанда, тұтынылатын тағамның мөлшері мен сапасы және оның ағзаға ену ритағы, бұлшықет және жүйке қызметінің деңгейі және т. б.Жануар ағзасы үйлестіру механизмдерінің күрделі кешені арқылы тамақтану режимінің сипатына, жүйке немесе бұлшықет жүктемесіне бейімделеді. Сонымен, көмірсулар мен липидтер алмасуының әртүрлі реакцияларының барысын бақылау жасуша деңгейінде тиісті субстраттар мен
ферменттердің концентрациясымен, сондай-ақ белгілі бір реакция өнімдерінің жинақталу дәрежесімен жүзеге асырылады. Бұл бақылау механизмдері өзін-өзі реттеу механизмдеріне жатады және бір жасушалы және көп жасушалы организмдерде жүзеге асырылады. Соңғысында көмірсулар мен майларды жоюдың реттелуі жасушааралық өзара әрекеттесу деңгейінде болуы мүмкін. Атап айтқанда, метаболизмнің екі түрі де өзара бақыланады: бұлшық еттердегі ЭЖК глюкозаның ыдырауын тежейді, ал май тініндегі глюкозаның ыдырау өнімдері ЭЖК түзілуін тежейді. Ең жоғары ұйымдастырылған жануарларда бүкіл организмнің метаболикалық процестерін бақылауда маңызды болып табылатын эндокриндік жүйенің эволюциясы процесінде пайда болуымен анықталатын аралық алмасуды реттеудің арнайы жасушааралық механизмі пайда болады.Омыртқалы жануарлардағы май мен көмірсулар алмасуын реттеуге қатысатын гормондардың ішінде келесілер орталық орын алады: тағамның қорытылуын және ас қорыту өнімдерінің қанға сіңуін бақылайтын асқазан-ішек гормондары; инсулин және глюкагон — көмірсулар мен липидтердің аралық алмасуының арнайы реттегіштері; СТГ және онымен функционалды байланысты "соматомединдер" және СИФ,глюкокортикоидтар, ACTH және адреналин спецификалық емес бейімделу факторлары болып табылады. Айта кету керек, көптеген аталған гормондар ақуыз алмасуын реттеуге де тікелей қатысады (9-тарауды қараңыз). Аталған гормондардың секреция жылдамдығы және олардың тіндерге әсерін жүзеге асыру өзара байланысты.Біз шырын бөлудің жүйке-гуморальды фазасына бөлінетін асқазан-ішек жолдарының гормоналды факторларының жұмысына арнайы тоқтала алмаймыз. Олардың негізгі әсерлері адам мен жануарлардың жалпы
физиологиясы курсынан жақсы белгілі, сонымен қатар олар туралы ч.3-те толық айтылған. Көмірсулар мен майлардың аралық метаболизмінің
эндокриндік реттелуіне толығырақ тоқталайық.Гормондар және аралық көмірсулар алмасуын реттеу. Омыртқалы организмдегі көмірсулар алмасуының интегралды көрсеткіші қандағы глюкозаның концентрациясы болып табылады. Бұл көрсеткіш тұрақты және сүтқоректілерде шамамен 100 мг% (5 ммоль/л) құрайды. Оның ауытқулары әдетте ±30% - дан аспайды. Қандағы глюкозаның деңгейі, бір жағынан, моносахаридтің негізінен ішектен, бауырдан және бүйректен қанға түсуіне, екінші жағынан, оның жұмыс істейтін және сақтайтын тіндерге ағуына байланысты (сурет. 2).
Бауыр мен бүйректен глюкозаның ағымы бауырдағы гликоген фосфорилаза мен гликоген синтетаза реакциясының белсенділік қатынасымен, глюкозаның ыдырау қарқындылығы мен бауырдағы және ішінара бүйректегі глюконеогенез қарқындылығымен анықталады. Глюкозаның қанға енуі фосфорилаза реакциясы мен глюконеогенез процестерінің деңгейімен тікелей байланысты. Глюкозаның қаннан тінге ағуы оның бұлшықет, адипозды және лимфоидты жасушаларға тасымалдану жылдамдығына тікелей байланысты, олардың мембраналары оларға глюкозаның енуіне кедергі жасайды (бауыр, ми және бүйрек жасушаларының мембраналары моносахаридке оңай өтетінін еске түсіріңіз); глюкозаның метаболикалық кәдеге жаратылуы, өз кезегінде оған өткізгіштікке тәуелді және оның ыдырауының негізгі ферменттерінің белсенділігінен; глюкозаның бауыр жасушаларында гликогенге айналуы (Левин және басқалар, 1955; Ньюшолм, Рэндл, 1964; Фоа, 1972). Глюкозаның тасымалдануы
нейросекреторлық ядролар мен орталықтар локализацияланған. Гипоталамус-гипофизді реттеу кері байланыс принципі бойынша жұмыс істейтін механизмдер арқылы жүзеге асырылады, онда өзара әрекеттесудің әртүрлі деңгейлері айқын ерекшеленеді (сурет. 1).
Сурет 2. Керібайланысдеңгейлері.
"Ұзын" керібайланыс тізбегі перифериялық эндокриндік бездің гипофиз және гипоталамус орталықтарымен (ОЖЖ-ніңсупрагипоталамикалық және басқа аймақтарымен)әсер ету " керібайланыс тізбегі гипофиздің тропиктік гормонының жоғарылауы (мысалы, ACTH) гипофизотропты гормонның (бұл жағдайда кортиколиберин) секрециясы мен бөлінуін модуляциялайты және өзгертетін өзара әрекеттесуді білдіреді."Ультра қысқа" керібайланыстізбегі- біргипофизотропты гормонның бөлінуі басқа гипофизотропты гормонның бөлінуі мен бөліну процестеріне әсер ететін гипоталамус ішіндегі өзара әрекеттесутүрі. Кері байланыстың бұл түрікез- келген эндокриндік безде болады. Сонымен, осы нейрондардың аксондары арқылы және жасуша аралық өзара әрекеттесу арқылы
(жасушадан жасушаға) окситоцин немесе вазопрессиннің бөлінуі осы гормондарды шығаратын нейрондардың белсенділігін өзгертеді. Тағы бірмысал, пролактиннің бөлінуі және оның васкулярлық кеңістіктерге диффузиясык өршілес лактотрофтарға әсер етеді, соданкейін пролактин секрециясының тежелуі."Ұзын "және" қысқа "керібайланыс тізбектері" жабық " типті жүйелер ретінде жұмыс істейді, яғни олар өзін-өзі реттейтін жүйелер.Алайда, олар ішкі және
сыртқы сигналдарға жауап береді, қысқа уақытқа өзін- өзі реттеу принципін өзгертеді (мысалы, стресс кезінде және т.б.). Сонымен қатар,бұл жүйелерге күн мен түннің өзгеруіне байланысты биологиялық циркадиялық ритақты қолдайтын механизмдер әсер етеді. Күндізгі-түнгі меақтуралы ақпарат Күн-түн механизмінен басқа, осы "сағаттардың" қызметіне басқа реттеушілер қатысады (Дене температурасының өзгеруі, демалу, ұйқы және т.б.).
2.2. Көмірсу, липид, ақуыз және су - тұз алмасуының гормоналды реттелуі Тірі организмнің негізгі энергетикалық ресурстары — көмірсулар мен
майлар ферменттік катаболикалық түрлендірулер арқылы жасушаларда оңай алынатын потенциалдық энергияның жоғарық орына ие. Көмірсулар мен май алмасуларының, сондай-ақ гликолиз өнімдерінің биологиялық тотығу процесінде бөлінетін Энергия негізінен синтезделген АТФ фосфат байланыстарының химиялық энергиясына айналады.АТФ-да жинақталған макроэргиялық байланыстардың химиялық энергиясы, өз кезегінде, әртүрлі жасушалық жұмыстарға жұмсалады. — Электрохимиялық градиенттерді құружәне қолдау, бұлшық еттердің жиырылуы, секреторлық және кейбір көлік процестері, ақуыз, май қышқылдарының биосинтезі және т. б. "отын" функциясынан басқа, көмірсулар мен майлар ақуыздармен бірге маңызды құрылыс, пластикалық жеткізушілер рөлін атқарады. Жасушаның негізгі құрылымдарына кіретін материалдар-нуклеин қышқылдары, қарапайым ақуыздар, гликопротеидтер, бірқатар липидтер және т.б. Көмірсулар мен майлардың ыдырауына байланысты синтезделген
АТФ жасушаларды жұмыс
істеугеқажетті энергиямен қамтамасыз етіп қана қоймайды, сонымен қатар camp түзілу көзі болып табылады, сонымен қатар көптеген ферменттердің белсенділігін, құрылымдық ақуыздардың күйін реттеуге қатысады, олардың фосфорлануын қамтамасыз етеді.Жасушалар тікелей тастайтын көмірсулар мен липидті субстраттар-моносахаридтер (ең алдымен глюкоза) және эфирленбеген май қышқылдары (HEFA), сондай-ақ кейбір тіндерде кетон денелері. Олардың көздері-көмірсулардың гликоген түрінде және бейтарап майлар түрінде ағзаларға салынған ішектен сіңетін Тамақ өнімдері
липидтер, сондай-ақ көмірсулар түзетін аминқышқылдары мен глицерин (глюконеогенез). Омыртқалылардағы депозиттік органдарға бауыр мен майлы (адипозды) тін, глюконеогенез органдарына — бауыр мен бүйрек жатады. Жәндіктерде май ағзасы депозиттік орган болып табылады. Сонымен қатар, глюкоза мен HEHA көздері жұмыс істейтін жасушада сақталған немесе өндірілген кейбір қосалқы немесе басқа өнімдер болуы мүмкін. Көмірсулар мен май алмасуының әртүрлі жолдары мен кезеңдері көптеген өзара әсерлермен байланысты. Бұл алмасу процестерінің бағыты мен қарқындылығы бірқатар сыртқы және ішкі факторларға байланысты. Оларға, атап айтқанда, тұтынылатын тағамның мөлшері мен сапасы және оның ағзаға ену ритағы, бұлшықет және жүйке қызметінің деңгейі және т. б.Жануар ағзасы үйлестіру механизмдерінің күрделі кешені арқылы тамақтану режимінің сипатына, жүйке немесе бұлшықет жүктемесіне бейімделеді. Сонымен, көмірсулар мен липидтер алмасуының әртүрлі реакцияларының барысын бақылау жасуша деңгейінде тиісті субстраттар мен
ферменттердің концентрациясымен, сондай-ақ белгілі бір реакция өнімдерінің жинақталу дәрежесімен жүзеге асырылады. Бұл бақылау механизмдері өзін-өзі реттеу механизмдеріне жатады және бір жасушалы және көп жасушалы организмдерде жүзеге асырылады. Соңғысында көмірсулар мен майларды жоюдың реттелуі жасушааралық өзара әрекеттесу деңгейінде болуы мүмкін. Атап айтқанда, метаболизмнің екі түрі де өзара бақыланады: бұлшық еттердегі ЭЖК глюкозаның ыдырауын тежейді, ал май тініндегі глюкозаның ыдырау өнімдері ЭЖК түзілуін тежейді. Ең жоғары ұйымдастырылған жануарларда бүкіл организмнің метаболикалық процестерін бақылауда маңызды болып табылатын эндокриндік жүйенің эволюциясы процесінде пайда болуымен анықталатын аралық алмасуды реттеудің арнайы жасушааралық механизмі пайда болады.Омыртқалы жануарлардағы май мен көмірсулар алмасуын реттеуге қатысатын гормондардың ішінде келесілер орталық орын алады: тағамның қорытылуын және ас қорыту өнімдерінің қанға сіңуін бақылайтын асқазан-ішек гормондары; инсулин және глюкагон — көмірсулар мен липидтердің аралық алмасуының арнайы реттегіштері; СТГ және онымен функционалды байланысты "соматомединдер" және СИФ,глюкокортикоидтар, ACTH және адреналин спецификалық емес бейімделу факторлары болып табылады. Айта кету керек, көптеген аталған гормондар ақуыз алмасуын реттеуге де тікелей қатысады (9-тарауды қараңыз). Аталған гормондардың секреция жылдамдығы және олардың тіндерге әсерін жүзеге асыру өзара байланысты.Біз шырын бөлудің жүйке-гуморальды фазасына бөлінетін асқазан-ішек жолдарының гормоналды факторларының жұмысына арнайы тоқтала алмаймыз. Олардың негізгі әсерлері адам мен жануарлардың жалпы
физиологиясы курсынан жақсы белгілі, сонымен қатар олар туралы ч.3-те толық айтылған. Көмірсулар мен майлардың аралық метаболизмінің
эндокриндік реттелуіне толығырақ тоқталайық.Гормондар және аралық көмірсулар алмасуын реттеу. Омыртқалы организмдегі көмірсулар алмасуының интегралды көрсеткіші қандағы глюкозаның концентрациясы болып табылады. Бұл көрсеткіш тұрақты және сүтқоректілерде шамамен 100 мг% (5 ммоль/л) құрайды. Оның ауытқулары әдетте ±30% - дан аспайды. Қандағы глюкозаның деңгейі, бір жағынан, моносахаридтің негізінен ішектен, бауырдан және бүйректен қанға түсуіне, екінші жағынан, оның жұмыс істейтін және сақтайтын тіндерге ағуына байланысты (сурет. 2).
Бауыр мен бүйректен глюкозаның ағымы бауырдағы гликоген фосфорилаза мен гликоген синтетаза реакциясының белсенділік қатынасымен, глюкозаның ыдырау қарқындылығы мен бауырдағы және ішінара бүйректегі глюконеогенез қарқындылығымен анықталады. Глюкозаның қанға енуі фосфорилаза реакциясы мен глюконеогенез процестерінің деңгейімен тікелей байланысты. Глюкозаның қаннан тінге ағуы оның бұлшықет, адипозды және лимфоидты жасушаларға тасымалдану жылдамдығына тікелей байланысты, олардың мембраналары оларға глюкозаның енуіне кедергі жасайды (бауыр, ми және бүйрек жасушаларының мембраналары моносахаридке оңай өтетінін еске түсіріңіз); глюкозаның метаболикалық кәдеге жаратылуы, өз кезегінде оған өткізгіштікке тәуелді және оның ыдырауының негізгі ферменттерінің белсенділігінен; глюкозаның бауыр жасушаларында гликогенге айналуы (Левин және басқалар, 1955; Ньюшолм, Рэндл, 1964; Фоа, 1972). Глюкозаның тасымалдануы