Файл: ан айналымы биохимиясы модулі бойынша апаратты топтама.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 12.12.2023
Просмотров: 344
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
СОДЕРЖАНИЕ
ЛИПОПРОТЕИНДЕР ЖӘНЕ ЛИПИДТЕРДІҢ ТАСЫМАЛДАНУЫ
(см. электив «Кардиомаркеры»)
А. Структура и функции липопротеинов (см. табл.1).
Липидтер әртүрлі функцияларды орындайды. Олар жасуша мембраналарының құрамына кіреді, стероидты гормондардың, өт қышқылдарының, простагландиндердің және фосфоинозитидтердің бастамалары ретінде қызмет етеді. Қанда липидтердің жеке компоненттері (қаныққан, моно-және полиқанықпаған май қышқылдары), триглицеридтер, холестерин, холестерин эфирлері және фосфолипидтер бар. Барлық осы заттар суда ерімейді, сондықтан ағзада липидтерді тасымалдаудың күрделі жүйесі бар. Бос (этерификацияланбаған) май қышқылдары альбуминмен кешендер түрінде қанға тасымалданады. Триглицеридтер, холестерин, холестерин эфирлері және фосфолипидтер суда еритін липопротеидтер түрінде тасымалданады. Липопротеиндер – бұл гидрофобты өзек пен гидрофильді қабықтан тұратын сфералық бөлшектер. Өзек құрамында полярлы емес липидтер – триглицеридтер және холестерин эфирлері бар. Қабық полярлы липидтерден – холестериннен және фосфолипидтерден тұрады, осы молекулалардың зарядталған ұштары сыртқа қарайды. Сонымен қатар, қабықтың құрамына фосфолипидтер мен холестеринмен ковалентті емес байланысқан белоктар – апопротеиндер кіреді. Апопротеиндер липопротеидті бөлшектердің құрылымын сақтайды және олардың липопротеидті рецепторлармен әрекеттесуін қамтамасыз етеді. Қан айналымында липопротеидті бөлшектер бір-бірімен үстіңгі липидтер мен апопротеиндер алмасады. Апопротеиндер липопротеиндердің "визит карточкасы" ретінде қызмет етеді, өйткені әртүрлі жасушалардағы липопротеиндердің рецепторлары тек белгілі бір апопротеиндерді таниды.
Липопротеидті бөлшектердің тығыздығы апопротеин / липидтердің қатынасы арқылы анықталады: бұл қатынас неғұрлым үлкен болса, тығыздық соғұрлым жоғары болады. Электрофорезде жылжуы апопротеиндер мен полярлы липидтердің мөлшерінен тәуелді.
Липопротеиндерді олардың тығыздығына (ультрацентрифугалау арқылы бағаланады) және электрофорез кезінде жылжуына байланысты бірнеше кластарға бөледі. Қан плазмасының липопротеин спектрін анықтау медицинада атеросклерозды диагностикалау үшін қолданылады.
Қандағы әртүрлі тығыздықтағы липопротеиндердің құрамын анықтау кезінде оларды әдетте электрофорез әдісімен бөледі. Бұл жағдайда ХМ стартта қалады (γ-глобулиндерге ұқсас), β-ЛП, пре-β-ЛП және α-ЛП, сәйкесінше β-, α1 және α2 глобулиндердің орнын алады. Егер қанда β-глобулиндердің (ТТЛП) мөлшері жоғары болса, бұл холестериннің тіндерге жиналуын білдіреді (атеросклероз дамиды).
Аш қарынға алынған қанда тек ТӨТЛП, ТТЛП және ТЖЛП болады, ал басқа липопротеиндер (хиломикрондар, хиломикрондардың қалдық компоненттері, сондай-ақ ТАЛП) тамақтан кейін немесе липидтер алмасуының бұзылуында ғана анықталады.
ХМ және ТӨТЛП-ң түзілуі
Хиломикрон (от «chyle» - сүт шырыны, лимфа; «micros» - кішкентай).
Хиломикрондар – экзогенді липидтердің (триглицеридтер мен холестериндердің) негізгі тасымалдаушы формасы.
Бұл кішкентай май тамшысы: оның ортасында триацилглицериндер болады, олар бөлшектің басым бөлігі болып табылады және хиломикрон массасының 80% құрайды. Шеткі жағында фосфолипидтер қабаттары (массаның 8%) және апобелоктар қабаттары (массаның 2%) орналасқан, олардың екеуі - А және В48 энтероцит рибосомаларында синтезделеді. Қалған 10% массасы холестерин мен оның эфирлеріне тиесілі. Хиломикронның беті гидрофильді: ақуыздар мен фосфолипидтердің гидрофильді бөліктері бөлшек бетінде орналасқан.
Майлы тағамдарды пайдаланғаннан кейін қанда хиломикрондар мөлшерінің жоғарылайтындығы байқалады.
Ұзын тізбекті МҚ бар ТАГ ағзаға тамақпен бірге түскен кезде ХМ пайда болады. Егер құрамында көміртегі атомдарының саны 12-ден аз МҚ бар ТАГ тағам арқылы ағзаға түсетін болса (олар әсіресе сүттің ТАГ-не бай), онда ішек қабырғасында ХМ түзілмейді, өйткені мұндай МҚ ішек қабырғасында сіңгеннен кейін лимфа жүйесін айналып өтіп, тікелей бауырға қақпа венасы арқылы енеді.
Хиломикрондар шырышты ішектің эпителий жасушаларында түзіледі.
Хиломикрондар көлемі үлкен болғандықтан экзоцитоз жолымен қан жүретін капиллярлар қабырғаларында болатын саңылаулар арқылы өте алмайды (диаметрі 100 ден 1000 нм дейін). Сондықтан да хиломикрондар экзоцитоз жолымен лимфаға түседі.
Оң жақ жүрек пен өкпеге, содан соң қан айналымының үлкен айналымына ауысады. Лимфа хиломикрондарында B48 апопротеині және А апопротеинінің бірнеше түр тармақтары бар. Қанда хиломикрондар С апопротеині мен Е апопротеинінің бірнеше түр тармақтары бар ТЖЛП-мен кездеседі. Хилимикрондар ТЖЛП-ң апопротеиндерімен алмасады: А апопротеиндерінің бір бөлігін беріп С және Е апопротеиндерін алады. Май тіндері, миокард, қаңқа бұлшықеттері, сүт бездерінің қан жүретін капиллярларында хиломикрондар эндотелий беткейінде орналасқан липопротеидлипазамен ыдырайды. Липопротеидлипазаның кофакторы СII апопротеині болып табылады. Липопротеинлипаза хиломикронның триацилглицериндерін гидролиздейді. Сонымен қатар, олардың барлығы тамырішілік липолиздің есебінен айтарлықтай мөлшерде ТАГ жоғалтады және олардың гидрофобты ядросы айтарлықтай "салмағын жоғалтады". ХМ фосфолипидті қалдық бөліктерге айналады, ХС және апопротеидтер салыстырмалы түрде артық болады.
Қабықтың компоненттері ТЖЛП бөлшектерімен ұсталады, ал қаннан хиломикрондардың қалдық компоненттері бауырға шығарылады. Бауырда E апопротеин рецепторлары бар.
Хиломикрондардың өздері атерогендік қасиетке ие емес, бірақ хиломикрондардың қалдық компоненттері атерогенді болып табылады.
Ферменттердің дисфункциясы
ХМ – үлкен бөлшектер, жарықты шашыратады, сондықтан қандағы ХМ жоғары концентрациясында плазма бұлдыр (сүт түсті) болады.
Қан ағымынан ХМ-ң жойылу жылдамдығы төмендегілерге тәуелді:
-
ТЖЛП және С-II және Е құрылымдарының болуы; -
С-II және Е апопротеиндердің ТЖЛП арқылы ХМ-ға тасымалдану жылдамдығы; -
Липопротеинлпаза белсенділігі;
Осы ауысуларға қатысатын кез-келген белок құрылымының бұзылуы отбасылық гиперхиломикронемияға және I типті гипертриацилглицеролемияға әкеледі.
Липопротеидлипазаның шын жетіспеушілігі әдетте балалық шақта әртүрлі органдар мен тіндерде майлардың жиналуын көрсететін белгілермен анықталады: теріде (бөртпе ксантоматозымен бірге); бауырда (гепатомегалия); көздің тор қабығының қан тамырларында (ретинальды липемия); іштің ауыруы (гиперхиломикронемиямен бірге жүретінбелгі). Тамыр саңылауларының тарылуы, есте сақтау қабілетінің, ұйқы безі функцияларының бұзылуы байқалады.
С-II апопротеинінің жетіспеушілігіне негізделген хиломикронемия ересектерде жиі байқалады.
2. ТӨТЛП негізінен бауырда түзіледі. Бұл бауырда синтезделген эндогендік триглицеридтердің негізгі тасымалдаушы түрі. ТӨТЛП – қаңқа бұлшықеттері мен миокард, май тіндері үшін негізгі триглицеридтерді жеткізушісі. ТӨТЛП құрамына C, E және B100 апопротеиндері кіреді. Хиломикрондармен салыстырғанда ТӨТЛП құрамында триглицеридтер аз, бірақ холестерин мен холестерин эфирлері көп.
Триглицеридтер бауырда май тіндерінен түсетін БМҚ-нан синтезделеді. Рационда көмірсулардың жоғары деңгейі кезінде глюкозаның артық болуынан да БМҚ-ң тікелей синтезі болуы мүмкін. Бұл триглицеридтер, сондай-ақ, бауырда синтезделген немесе хиломикрондардың қалдықтарынан түскен холестерин В, Е және С апопротеиндерімен қосылып, ТӨТЛП түзеді. Бұл липопротеиндердің одан әрі метаболизмі жетілу кезеңін қамтиды, ол ТЖЛП-ден С-II және Е 2 апопротеиндердің ТӨТЛП-ге ауысуынан тұрады. Апо-С-II капиллярлардың қабырғаларында липопротеинлипазасын белсендіреді, ал хиломикрондардың экзогендік триглицеридтері секілді триглицеридтері гидролизге ұшырайды және ТОЛП қалатын қан плазмасынан жойылады. ТОЛП-ң кейбір бөлігі бауыр рецепторларымен өзара әрекеттеседі, ал қалғандары холестериннен және апопротеиннен тұратын ТТЛП-ге айналады. Бұл ауысу процесінің механизмі мен жиналуы белгісіз.
3. ТОЛП (ТАЛП) – тығыздығы орташа немесе аралық тығыздықты липопротеиндер. Қаңқа бұлшықеттерінің, миокардтың және май тіндерінің капиллярларында ТӨТЛП липопротеинлипазамен ыдырап, ТОЛП түзеді. ТОЛП құрамында E және B100 апопротеиндері бар. Бауырда ТОЛП-ң жартысы триацилглицеролипазаның әсерінен ТТЛП-ге айналады, ал қалған ТТЛП гепатоциттердің рецепторларымен байланысып, эндоцитозға ұшырайды және жойылады. IIB фенотипі бар отбасылық гиперхолестеринемия кезінде III типті гиперлипопротеинемия және майлы тағамдарды қолдану кезінде ТОЛП деңгейі жоғарылайды, бұл атеросклероз қаупін арттырады.
4. ТТЛП – тығыздығы төмен липопротеиндер.
ТТЛП-ң көп бөлігі, жоғарыда сипатталғандай, ТӨТЛП-ден түзіледі, LDL-нің бір бөлігі тікелей бауырда синтезделеді. ТӨТЛП мен ТОЛП ыдырауының соңғы өнімі ТТЛП болып табылады. ТТЛП - бұл ең атерогенді липопротеиндер. ТТЛП құрамында барлық плазмалық холестериннің үштен екісі, негізінен холестерин эфирлері түрінде болады.
Басқа липопротеиндерден айырмашылығы, ТТЛП-де тек B100 апопротеині бар.
ТТЛП-ң қызметі: бауырдан тіндерге холестеролдың тасымалдануы.
ТТЛП құрамындағы ХС-ң жасушаға рецепторлы-жанамаланған эндоцитоз арқылы тасымалдау механизмі 1985 жылы Нобель сыйлығымен марапатталған американдық ғалымдар M. Brown және J. Goldstein-дің керемет жұмыстарының арқасында белгілі.
Плазмадан ТТЛП-ді шығарудың 2 жолы бар.
ТТЛП-ң шамамен 70%-ы қаннан бауырға жоғары спецификалық ТТЛП рецепторлары – B, E - апопротеин рецепторлары арқылы шығарылады. Сурет 1.
Біріншісіне сәйкес, ТТЛП көптеген жасушалардың мембраналарының бетінде болатын арнайы рецепторлармен байланысқаннан кейін рецептор-тәуелді эндоцитоз арқылы жасушалармен ұсталынады. ТТЛП-рецептор кешені эндосомаға қосылады және лизосомамен бірігеді. Эндосомалардағы қышқыл орта ТТЛП-рецептор кешендерінің диссоциациясына ықпал етеді. Босатылған рецептор плазмалық мембранаға оралады. Лизосомалық ферменттер холестеролдан басқа ТТЛП-ң барлық компоненттерін гидролиздейді. Бұл жағдайда ЛП ақуызы аминқышқылдарына, ал ХСЭ – бос XС мен БМҚ дейін ыдырайды. ТАГ мен ФЛ де ыдырайды.
Сурет 1.
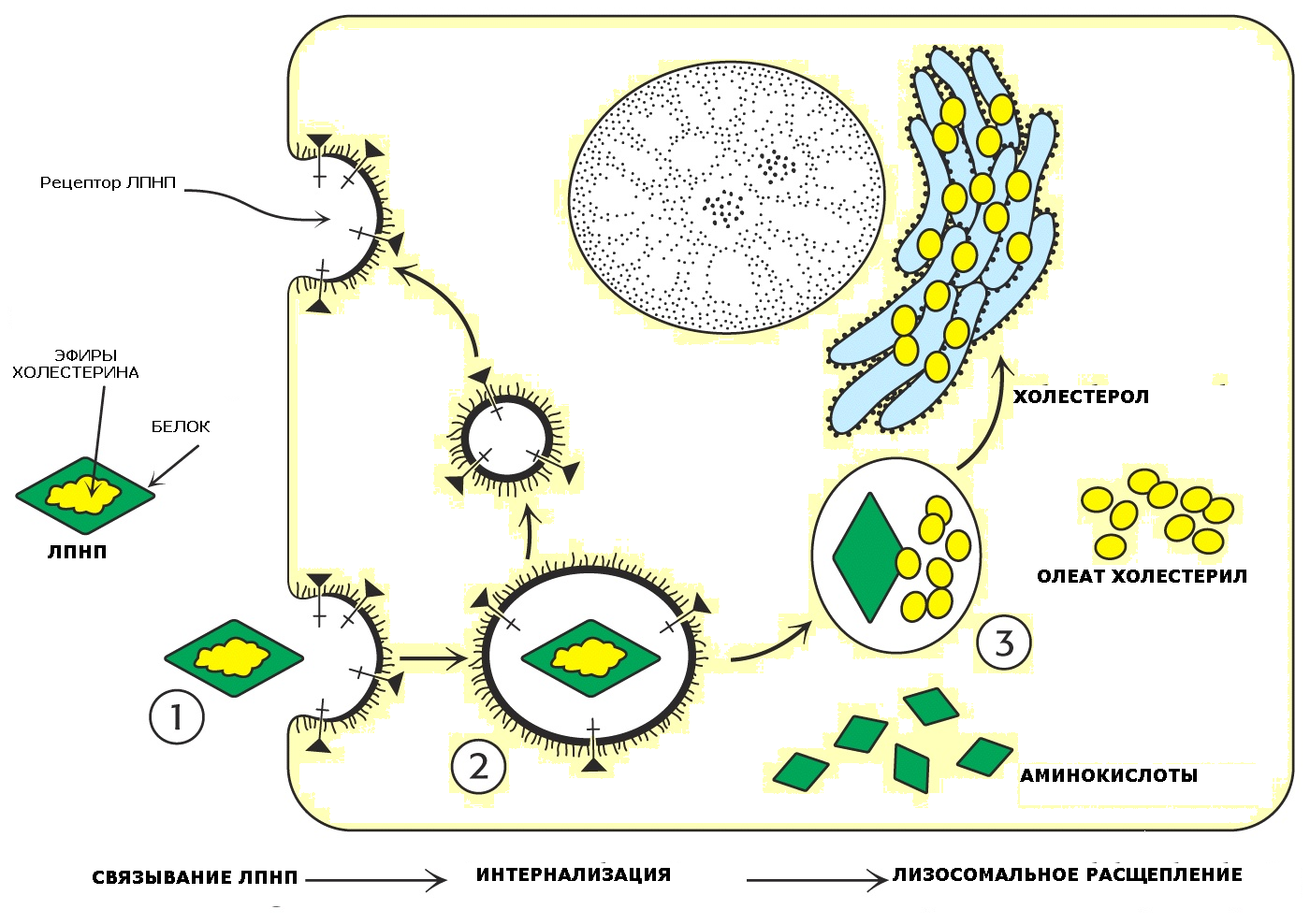
Лизосомадан босап шыққан холестерол қабілетті:
-
Биомембрана құрамына кіреді; -
Басқа стероидтар мен өт қышқылдарын синтездеу үшін пайдаланылады; -
Жасушалардағы холестериннің биосинтезін, сондай-ақ жасуша бетіндегі ТТЛП-ң В,Е- апопротеин рецепторларының биосинтезін тежейді, жасушаішілік холестерин деңгейін реттейді. ТТЛП рецепторларының синтезін реттеуге инсулин және Т3 гормондар, жыныс гормондары қатысады. Олар ТТЛП рецепторларының түзілуін арттырады, ал кортизол азайтады. -
Соңында, бос ХС микросомальді энзим – ацетил-КоА-холестерин-ацил-трансферазаны (АХАТ), эстерифициялаушы ХС-ді белсендіреді, осылайша цитоплазмада ХСЭ депонизациясына ықпал етеді. Осындай күрделі жолмен бір жағынан, жасушадағы XС синтезі, екінші жағынан, ТТЛП-ң жасушаға енуін және олармен XС жеткізілуі бақыланады. -
ТТЛП-ның қалған 30% аз спецификалық бейтараптандырушы рецепторлардың көмегімен ретикулоэндотелиалдық жүйе жасушалары ұстап алады. Бауырдағы B, E-апопротеин-рецепторлардың синтезі ТТЛП концентрациясы жоғарылағанда басылады. Керісінше, ретикулоэндотелий жүйесінің жасушаларында бейтараптандыратын рецепторлардың саны ТТЛП деңгейіне тәуелді емес.