Файл: ан айналымы биохимиясы модулі бойынша апаратты топтама.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 12.12.2023
Просмотров: 349
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
СОДЕРЖАНИЕ
Липидтерді және полиқанықпаған май қышқылдарын (ПҚМҚ) тасымалдау (элективке қосымша материал)
Қаныққан және полиенді МҚ-н жасушаларға бөлек тасымалдау кезінде, екеуі де және басқалары да жасушаларға пассивті және активті жолменде енеді (сурет 2). МҚ полярлы липидтерден жасушаға пассивті жолмен ЛП жасушаларының сіңіруінсіз түсетіндігі олардың маңызды айырмашылығы болып табылады. МҚ-ң белсенді тасымалдануы кезінде жасушаға полярлы емес липидтердің құрамдас бөлігі ретінде енеді, сондықтан жасушалар рецепторлық эндоцитоз арқылы ЛП-ді сіңіреді. Пассивті жол филогенетикалық тұрғыдан ежелгі; белсенді тасымалдау едәуір заманауи және едәуір жетілдірілген болып келеді.
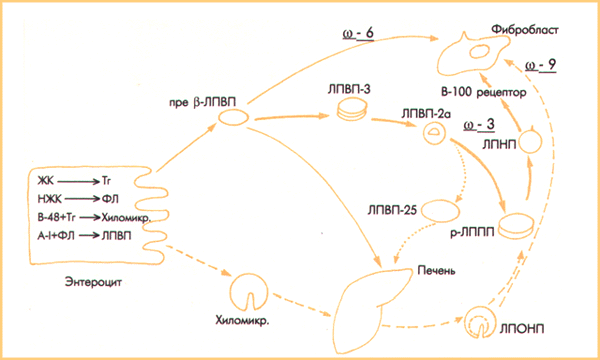
Сурет 2. Қаныққан және полиенді МҚ-н қанға липопротеиндермен тасымалдау.
Полиенді МҚ тасымалдау - тұтас сызықтар (жуан сызық –белсенді, жіңішке сызық - пассивті )
Холестерин эфирлеріндегі полиенді МҚ тасымалдау - нүкте сызығы.
Қаныққан МҚ тасымалдау – үзік сызықты сызық
Қаныққан МҚ пассивті жолмен жасушаға альбуминмен байланысу арқылы енеді; активті жолмен қаныққан МҚ жасушаға ТОЛП-ң апоЕ/В-100 рецепторлы эндоцитозы кезінде енеді. Пассивті түрде омега-6 полиенді МҚ тасымалдау мембраналық фосфолипидтер және ТЖЛП фосфолипидтері арасындағы МҚ-н трансэтерификациялау кезінде өтеді. Бірақ, омега-З МҚ жасушаға пассивті жолмен ене алмайды. Омега-З МҚ-на деген қажеттілік, борпылдақ дәнекер ұлпаларының жасушалары оларды эйкозаноидтар (простагландиндер, тромбоксандар, лейкотриендер) синтезі үшін қолданады. Эйкозаноидтар капиллярлардағы қан ағысын, тромбоциттердің агрегациясын, электролиттердің шығарылуын, қабыну реакциясын жергілікті реттейді. Арахидондық омега-6 және эйкозапентаендік Омега-З МҚ жасушасынан синтезделген эйкозаноидтар белсенділігі жағынан айтарлықтай ерекшеленеді. Эйкозаноидтар небәрі бірнеше минут ғана әсер етеді; олардың жасушалары сақталынбайды, сондықтан олардың синтезі мен әсер етуі бір уақытта жүреді. Бұл омега-З МҚ жасушаларына рецепторлық енуін қажет етеді. Бірақ, жасушаның рецепторлық эндоцитозымен тек қана полярлы емес күрделі липидтер енеді.
Бірінші кезеңде Омега-З МҚ жасушасына активті тасымалдау полярлы фосфолипидттерден полярсыз холестерин эфиріне (ХЭ) МҚ-н трансэтерификациялау болып табылады; реакция ТЖЛП-де жүреді. Екінші кезеңде холестерин эфирі (ХЭ) холестерин эфирін тасымалдаушы белок (ХЭТБ) әсері арқылы ТОЛП-ге ауысады. Гидрофильді холестерин эфирі (ХЭ) триглицеридттерді ауысытыра отырып, ТОЛП-дің ТТЛП-ге ауысуына көмектеседі. Егер қаныққан МҚ жасушаларына ТӨТЛП мен ТОЛП-ді тасымалданса, онда ТТЛП-ді Омега-З МҚ-на тән тек полиенділер тасымалдайды. Үшінші кезең – апоВ-100 эндоцитоз арқылы ТТЛП жасушаларын сіңіру болып табылады. АпоВ-100 эндоцитоз Омега-З МҚ-ң жасушаға енуінің негізгі кілті деп саналады. АпоВ-100 лиганд құрылымының генетикалық ақауы немесе апо В-100 рецепторларының мембранасында болмауы IIа типті гиперлипопротеинемияға (ГЛП) ұшыратады. Триглицеридтердің холестерин эфирлеріне толық ауыстырылмауынан және ТОЛП-ң ТТЛП-ге ауысуының бұзылуынан туындаған апоВ-100 лигандтың функционалдық ақауы IIб типті ГЛП-ң негізін құрайды(Сурет 3)
.
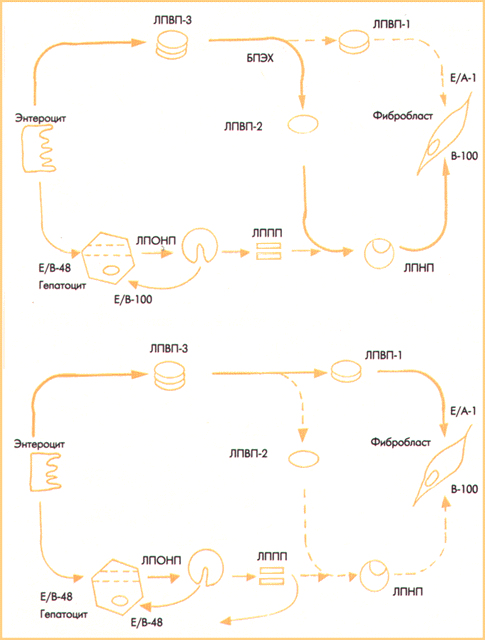
Сурет 3 – Полиенді МҚ жасушаларына тасымалдаудағы холестерин айналымы және Д.Фредриксон бойынша әртүрлі типтегі ГЛП-ның түзілуімен МҚ тасымалдау құрсауының орындары.
Лизосомаларда холестерин эфирінің гидролизінен жасушада қолданылатын полиенді МҚ босатылады. Холестерин цитозолдың тасымалдау ақуыздарын мембранаға өткізеді, ол арқылы қанға таралады. Қанда холестерин апоА-1У және ТЖЛП-ді байланыстырады, оны полиенді МҚ этерификациялау үшін қайта қолданады. Осылайша, полиенді МҚ-н тасымалдауда холестерин айналымы жүреді. Қандағы холестерин құрамын анықтай отырып, біз полиенді МҚ-ң тасымалдау формасы болып табылатын және жасушалар сіңіре алмайтын холестерин эфирі деңгейін бағалаймыз. Демек, қандағы холестерин полиенді МҚ жасушаларының жетіспеушілігін көрсетеді.
Алайда, apoВ-100 рецепторы полиенді МҚ-н тасымалдауда тек сүтқоректілердің бір бөлігінде (мұхит шошқалары, шошқалар, қояндар, приматтар), соның ішінде адамдарда негізгі кезең болып табылады; олардың барлығы қанда ХЭТБ-ң жоғары деңгейіне ие. Ал жануарлар (тышқандар, егеуқұйрықтар, иттер) қанында осы белоктың ізі ғана бар, холестерин эфирі ТЖЛП-тен ТТЛП-ге өтпейді және полиенді МҚ-н жасушаға тасымалдау apoВ-100 рецепторларының қатысуынсыз өтеді. Осы жануарларда холестерин эфирі ТЖЛП-тен ТТЛП-ге емес, ТӨТЛП-ге ауысады (апоА-1 + апоЕ белокты-липидті кешен (БЛК)), жасушалар апоЕ/А-1 эндоцитоз жолымен ТӨЖЛП-ді белсенді түрде сіңіреді. АпоЕ/А-1 рецепторлары арқылы полиенді МҚ-н активті тасымалдау филогенетикалық тұрғыдан ежелгі және эндогендік құрсауға төзімді болып келеді. ApoВ-100 эндоцитоз жолымен полиенді МҚ жасушаларына тасымалдануы жануарлардың атеросклерозға сезімталдығын анықтаса, ал apoЕ/А-1 эндоцитоз атеросклерозға төзімділілігін анықтайтындығын болжауға болады (сурет 4).
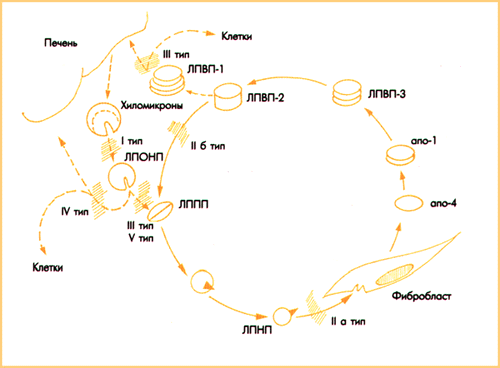
Сурет 4. Сүтқоректілердің атеросклерозға сезімталдығын (жоғарғы) және төзімділігін (төменгі) анықтайтын полиенді МҚ жасушаларына активті тасымалдаудың екі нұсқасы.
"Кардиомаркерлер" элективті топтамасы (жалғасы)
Атеросклерозды, полиенді МҚ жасушаларындағы тапшылық синдромы деп білеміз. Атеросклероздың симптомдары – ГЛП, гипертония, тромбоциттердің гиперагрегациясы, гиперинсулинемия және 11 типті қант диабеті, гиперкоагуляция, иммундық жүйенің гуморальды және жасушалық байланыстарының белсенділігі және қабыну. МҚ жасушаларындағы тасымалдаудың маңызды айырмашылығы патологияның екі түрін ажыратуға негіз береді – полиенді тасымалдаудың патологиясы және қаныққан МҚ-н тасымалдаудың патологиясы. Біріншісіне атеросклероз, екіншісіне-эссенциальды гипертония жатады.
Кесте 2. Апобелоктардың сипаттамасы
| Белок | Негізгі липопротеин тасымалдаушысы | Қызметі | Атерогенезге әсері |
| A-l | ТЖЛП | ТЖЛП-ң құрылымдық компоненті, ЛХАТ-ны белсендіреді, A-l рецептор арқылы жасушалық мембраналармен өзара байланысады | A-l қауіп |
| A-ll | ТЖЛП | ТЖЛП-ң құрылымдық компоненті, Бауырда липазаны белсендіреді? | анықталмаған |
| A-IV | Хиломмкрондар, ТЖЛП | ЛХАТ-ны белсендіреді, Холестериннің кері тасымалдаунына қатысуы мүмкін | анықталмаған |
| (а) | Лп(а) | Белгісіз | (а) қауіп |
| B48 | Хиломикрондар | Ішектен триглицеридтерді тасымалдау | анықталмаған |
| B100 | ЛПОНП, ЛПпП, ЛПНП | В/Е рецепторлары арықлы холестеринді жасушаға ендіру. Бауырдан триглицеридтерді тасымалдау | B100 қауіп |
| C-I | Хиломикрондар, ТӨТЛП, ТЖЛП | in vitro жағдайында ЛХАТ-ны белсендіреді | анықталмаған |
| C-II | Хиломикроны, ЛПОНП, ЛПВП | Липопротеидлипазаны белсендіреді | анықталмаған |
| C-III | Хиломикрондар, ТӨТЛП, ТЖЛП | Триглицеридтермен байытылған ремнантты бөлшектердің бауырға сіңірілуін өзгерту. Липопротеидлипаза белсенділігін тежеп, ЛХАТ-ны белсендіреді. | C-III және A-l қауіп |
| D | ТЖЛП | Белгісіз | Анықталмаған |
| E | Хиломикрондар, ТӨТЛП, ТЖЛП-Е | В/Е рецепторлар арқылы холестериннің сіңірілуі, Бауырда ремнантов хиломикрондардың және ТӨТЛП-ң сіңірілуін қамтамасыз етеді. Эндотелиолды жасушаларменгепариноидтарды байланыстырады. Эфирмен байытылған холестериннің ТТЛП-бөлшектерінің түзілуіне және липидтердің тіндерде таралуына қатысады. Жүйке жүйесінің өсуі мен репарациясына әсер етеді. | E 3/4 и Е4/4 қауіп |
| H | Хиломикрондар | Прокоагулянтты белсенділікті реттейді | |
Липидтер алмасуының бұзылуы. Модификацияланған липопротеиндер және атеросклероз дамуының жасушалық механизмдері.
Гиперхолестеролемия гиперлипопротеинемияның IIа, IIб, III сияқты атерогенді типтерінде байқалады. Бұл тәждік атеросклероздың қауіптілік факторы болып табылады; ол жүректің ишемиялық ауруы мен миокард инфарктісінің клиникалық көріністерінің жиілігімен байланысты.
Бірқатар патологиялық жағдайларда екіншілік гиперхолестеролемия дамиды. Ол бауыр ауруларында бауыр ішілік және бауырдан тыс холестаз, бүйрек зақымдануы, ұйқы безі мен простата бездерінің қатерлі ісіктері, подагра, гипертониялық ауру, созылмалы алкоголизм, семіздік түрінде кездеседі.
Қан сарысуындағы оның күрт өсуі (26 ммоль / л дейін) қант диабеті және липоидты нефрозы бар науқастарда байқалады.
Аштық кезінде, орталық жүйке жүйесі зақымдануы кезінде, жедел жұқпалы аурулар, жедел панкреатит, безгек, пневмония, анемия, гемолитикалық сарғаю, жедел гепатит, бауыр мен ішектің қатерлі ісіктері, ревматизм, альфа-липопротеин жетіспеушілігі, абеталипопротеинемия кезінде гипохолестеролемия байқалады.
Гипохолестеролемия онкопатологияның пайда болуына жиі жағдай жасайды деп саналады (немесе онымен бірге жүреді).
Қан плазмасындағы холестериннің фракциялық құрамын және оның жеке липопротеидтерін (бірінші кезекте ТЖЛП) анықтау үлкен диагностикалық мәнге ие болды. Гепатоциттерде түзілетін альбумин плазмадағы холестеринді эфирлеу жүйесінің арнайы емес активаторы ретінде қызмет етеді.
Бауырда синтезделетін ЛХАТ ферментінің (лецитин-холестерол-ацилтрансфераза) арқасында бос холестерин ТЖЛП-ге эфирленеді. Бұл процесс қан плазмасында жүретіндігіне қарамастан, ол бауырдың функционалдық жетіспеушілігін көрсетеді. Әдетте қалыпты жағдайда холестериннің этерификация коэффициенті (яғни холестерин эфирінің жалпыға қатынасы) 0,6-0,8 (немесе 60-80%) құрайды, ал жедел гепатит пен созылмалы гепатиттің өршуінде, бауыр циррозында, обструктивті сарғаюда, созылмалы алкоголизм кезінде ол азаяды.
Атеросклероз патогенезі, клиникасы, диагностикасы, емі.
Қазіргі уақытта бұл аурудың пайда болуының бірыңғай теориясы жоқ. Бірақ келесі нұсқалар, сондай-ақ олардың тіркесулері ұсынылады:
-
липопротеидтердің инфильтрация теориясы - бірінші кезекте липопротеидтердің қан тамыр қабырғасында жинақталуы, -
эндотелий дисфункциясының теориясы - ең алдымен эндотелий мен оның медиаторларының қорғаныш қасиеттерінің бұзылуы, -
аутоиммундық - бірінші кезекте макрофагтар мен лейкоциттердің дисфункциясы, олардың қан тамыр қабырғасына енуі, -
моноклоналдық – бірыңғай салалы бұлшықет жасушаларының патологиялық клонының пайда болуы, -
вирустық - эндотелийдің алғашқы вирустық зақымдануы (герпес, цитомегаловирус және т.б.), -
пероксидтік - бірінші кезекте антиоксидантты жүйенің бұзылуы, -
генетикалық – қан тамыр қабырғасының бастапқы тұқым қуалайтын ақауы, -
хламидиалдық - қан тамыр қабырғасының хламидиямен біріншілік зақымдануы, негізінен хламидиапневмония (Chlamydiapneumoniae). -
гормондық - жасқа байланысты гонадотропты және адренокортикотропты гормондар деңгейінің жоғарылауы, холестерин үшін құрылыс материалы болып табылатын гормондар синтезінің жоғарылауына әкеледі.
Атеросклероз - қан тамырларының ауруы. Бұл жүйелі ауру, яғни бүкіл организмге әсер етеді. Аурудың мәніне келетін болсақ, артериялардың ішкі беткейінде (әдетте өте тегіс, тегіс және жылтыр) ерекше өсінділер - түйіндер пайда болады, олар дөңгелек немесе сопақша түзілімдер болып табылады, әдетте олардың беті тегіс емес және қан тамырларының қуысына қарап тұрады, осылайша, оның диаметрін азайтып, тамыр арқылы қан ағымына кедергі келтіреді.
Ересек адамның ағзасында шамамен 140 г холестерол бар (оның 130 г тіндердің холестеролы): 80% -ы жасуша мембраналарының бос холестерині және 10% -ы жасуша ішіндегі эфирленген холестерин. Қалыпты жағдайда холестериннің көп бөлігі (1,0 г дейін) организмде түзіледі (оның синтезі барлық мүшелер мен тіндердің жасушаларында жүреді) және 0,5 г дейін адам ағзасына тамақпен бірге түседі. Осылайша, 0,3 г холестеролды тамақпен бірге тұтыну оның қандағы концентрациясының 10-15% -ға ғана артуына әкеледі.
Бір күнде адам ағзасында 1,0 г-ға дейін холестерин жаңарады: ол бауыр жасушаларында (өт қышқылдары) тотықтырылады және сол мөлшерде өтпен, содан кейін нәжіспен немесе тері майымен шығарылады. Көп мөлшерде холестерин бауырда түзіледі (80%) - эндогенді холестериннің негізгі жеткізушісі, аш ішектің қабырғалары (15%) және теріде (5%). Бос холестерин жаңа жасушалық мембраналардың (әсіресе орталық жүйке жүйесінің жасушаларында көп холестерин), стероидты гормондардың, D дәрумені мен өт қышқылдарының түзілуіне жұмсалады.