Файл: Курс лекций, нгу 2000. Лекция Что такое молекулярная биология. Доказательства роли нуклеиновых кислот.doc
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 11.01.2024
Просмотров: 157
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
7. Помехоустойчивость.
Определение: Мутации замен нуклеотидов, не приводящие к смене класса кодируемой аминокислоты, называют консервативными. Мутации замен нуклеотидов, приводящие к смене класса кодируемой аминокислоты, называют радикальными.
В каждом триплете можно провести 9 однократных замен. Общее количество возможных замен нуклеотидов - 61 по 9 = 549. Из них:
23 замены нуклеотидов приводят к появлению кодонов - терминаторов трансляции.
134 замены не меняют кодируемую аминокислоту.
230 замен не меняют класс кодируемой аминокислоты.
162 замены приводят к смене класса аминокислоты, т.е. являются радикальными.
Из 183 замен 3-его нуклеотида, 7 приводят к появлению терминаторов трансляции, а 176 - консервативны.
Из 183 замен 1-ого нуклеотида, 9 приводят к появлению терминаторов, 114 - консервативны и 60 - радикальны.
Из 183 замен 2-го нуклеотида, 7 приводят к появлению терминаторов, 74 - консервативны, 102 - радикальны.
Итак: 364/162=2.25 (отношение числа консервативных замен к числу радикальных замен) - показатель помехоустойчивости генетического кода.
8. Неперекрываемость.
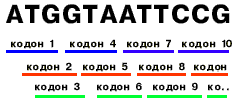 | В 1956г. Георгий Гамов предложил вариант перекрываемого кода. Согласно Гамовскому коду, каждый нуклеотид, начиная с третьего в гене, входит в состав 3-х кодонов. Когда генетический код был расшифрован, оказалось, что он неперекрываем, т.е. каждый нуклеотид входит в состав лишь одного кодона. |
Достоинства перекрываемого генетического кода:
компактность, меньшая зависимость структуры белка от вставки или делеции нуклеотида.
Недостаток:
большая зависимость структуры белка от замены нуклеотида и ограничение на соседей.
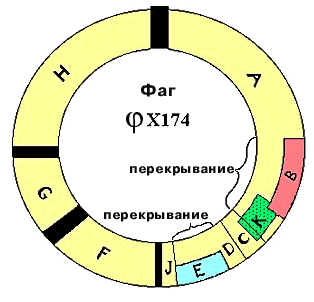 | В 1976г. была секвенирована ДНК фага φХ174. У него одноцепочечная кольцевая ДНК, состоящая из 5375 нуклеотидов. Было известно, что фаг кодирует 9 белков. Для 6 из них были определены гены, располагающиеся друг за другом. Выяснилось, что есть перекрывание. Ген Е полностью находится внутри гена D. Его инициирующий кодон появляется в результате сдвига считывания на один нуклеотид. Ген J начинается там, где кончается ген D. Инициирующий кодон гена J перекрывается с терминирующим кодоном гена D в результате сдвига на два нуклеотида. Конструкция называется "сдвиг рамки считывания" на число нуклеотидов, некратное трем. На сегодняшний день перекрывание показано только для нескольких фагов. |
Информационная емкость ДНК
На Земле живет 6 миллиардов человек. Наследственная информация о них заключена в 6х109 сперматозоидах. По разным оценкам у человека от 30 до 50 тысяч генов. У всех людей 30х1013 генов или 30х1016 пар нуклеотидов, которые составляют 1017 кодонов. Средняя книжная страница содержит 25х102 знаков. ДНК 6х109 сперматозоидов содержит информацию, равную по объему примерно 4х1013 книжных страниц. Эти страницы заняли бы объем 6-и зданий НГУ. 6х109 сперматозоидов занимают половину наперстка. Их ДНК занимает менее четверти наперстка.
Определение: транскрипция - это синтез всех видов РНК по матрице ДНК, осуществляемый ферментом ДНК-зависимой РНК-полимеразой.
У прокариот синтез всех видов РНК осуществляется одним и тем же ферментом.
У эукариот - 3 ядерные РНК-полимеразы, митохондриальные РНК-полимеразы, хлоропластные РНК-полимеразы.
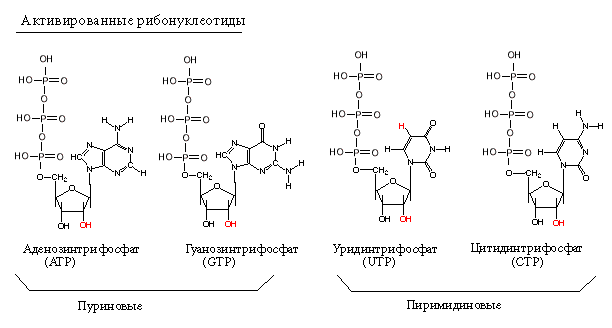
Субстратами для РНК-полимераз служат рибонуклеозид-трифосфаты (активированные нуклеотиды). Весь процесс транскрипции осуществляется за счет энергии макроэргических связей актвированных нуклеотидов.
Принципы транскрипции:
1. Комплементарность.
2. Антипараллельность.
3. Униполярность.
4. Беззатравочность.
5. Асимметричность.
РНК синтезируется комплементарно и антипараллельно транскрибируемой цепи ДНК. Рост цепи РНК идет только в направлении 5' 3'. Для начала синтеза РНК фермент не нуждается в поли- или олигонуклеотидной затравке.
Первый нуклеотид в РНК всегда пурин в форме трифосфата.
Понятие об опероне
Определение: оперон - единица транскрипции у прокариот.
В начале каждого оперона находится промотор.В конце каждого оперона находится терминатор. Перед терминатором располагаются структурные гены, или цистроны. Между промотором и цистронами может находиться оператор. Существуют моно-, олиго- и полицистронные опероны.
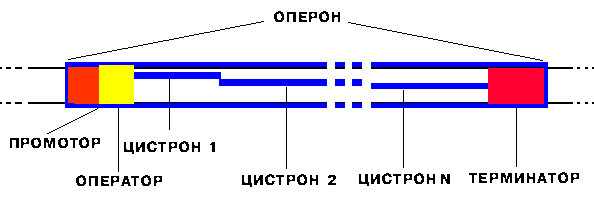
Определение: промотор - особая последовательность нуклеотидов ДНК, узнаваемая РНК-полимеразой как посадочная площадка и старт синтеза РНК.
Только с промотора может начаться синтез специфической РНК.
Определение: терминатор - особая последовательность нуклеотидов ДНК, узнаваемая РНК-полимеразой как финиш транскрипции.
Определение: цистрон - последовательность нуклеотидов ДНК, кодирующая один полипептид (в большинстве случаев - белок) или одну tРНК, или одну rРНК.
В большинстве случаев цистроны объединяются в оперон по следующему принципу: закодированные в них белки принимают участие в одной биохимической цепи реакций.
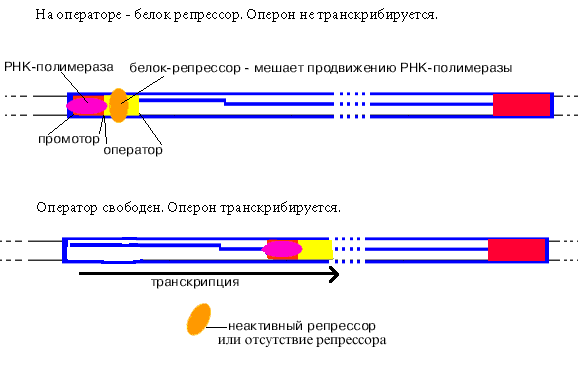
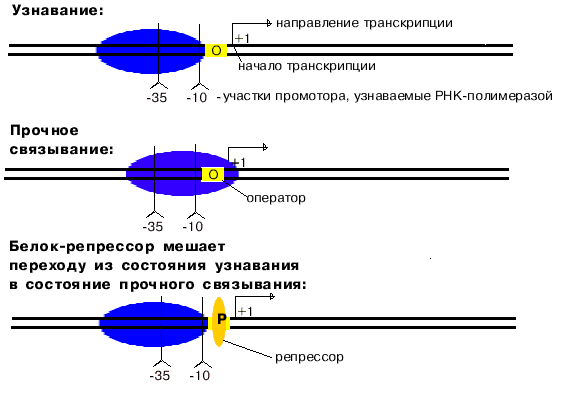
Определение: оператор - особая последовательность нуклеотидов ДНК, узнаваемая белком-репрессором.
У оператора диспетчерская функция - он разрешает или запрещает синтез РНК.
Асимметричность
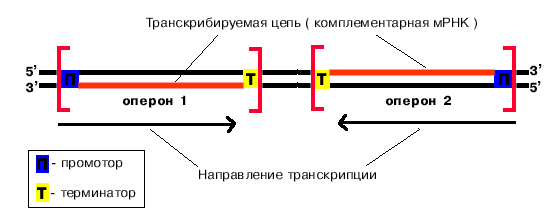
Транскрибируются обе цепи ДНК, но в каждом отдельном опероне только одна из них. Какая именно, определяется положением промотора и терминатора.
Субъединичный состав РНК-полимеразы Е.coli
РНК-полимераза Е.coli - белок с четвертичной структурой. Одновременно в клетке присутствует около 7000 молекул РНК-полимеразы.
Субъединичный состав фермента: (2 ) - holo-фермент (полный фермент). Без -фактора это core-фермент (2 ) .
(сигма) - фактор - сменный фактор специфичности.
Только holo-фермент обладает высоким сродством к специфической последовательности нуклеотидов - промотору, сродство к остальным случайным последовательностям ДНК у него снижено в 10000 раз. У core-фермента одинаковое сродство к любой последовательности нуклеотидов.
Сам по себе - фактор обладает наименьшим сродством к ДНК по сравнению с другими субьединицами РНК-полимеразы, однако он придает holo-ферменту такую конформацию, которая обладает повышенным сродством к промотору.
Как только произошла инициация транскрипции, -фактор отделяется. Элонгация - продолжение синтеза РНК, и терминация - его остановка, осуществляются core-ферментом.
Стадии узнавания и связывания, а также инициации осуществляются
holo-ферментом. Элонгация и терминация осуществляются core-ферментом.
Две субъединицы - каркас РНК-полимеразы. К ним крепятся остальные субъединицы.
- субъединица отвечает за прочное связывание с ДНК за счет кластера положительно заряженных аминокислот.
В - субъединице находятся два каталитических центра. Один отвечает за инициацию, а другой - за элонгацию. Один центр работает в holo-, а другой - в core- ферменте.
Особенности структуры промотора
Для изучения структуры промоторов провели следующий эксперимент. При оптимальных условиях связывания получили комплекс РНК-полимеразы с ДНК. Этот комплекс обработали ДНК-азой, и таким образом гидролизовали всю ДНК, незащищенную РНК-полимеразой. После этого отделили РНК-полимеразу от оставшихся фрагментов ДНК. Опять создали оптимальные условия для образования комплекса. Комплекс не образовывался.
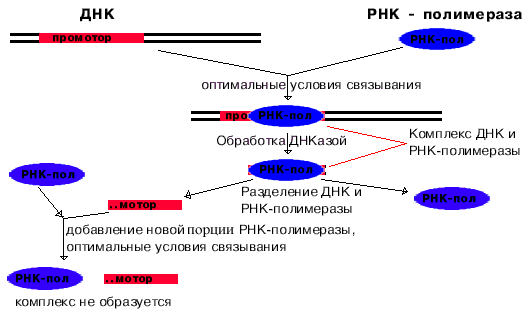
Отсюда следует вывод, что
узнавание и прочное связывание происходит на разных участках ДНК.
Эти участки отличаются и по первичной, и по вторичной структуре. Путем секвенирования выявили структуру многих промоторов. У большинства из них имеется общее свойство.
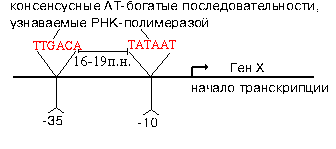
РНК-полимераза узнает промотор, покрывая 40-60 пар нуклеотидов. В промоторе узнается взаимное расположение двух расплавленных AT-богатых участков. В каждом из них расплавлено 4-6 пар. Центры этих участков находятся в положенях "-10" и "-35". Принципиально важным является расстояние между расплавленными участками. Оно колеблется от 16 до 19 п.н. Искусственное увеличение этого расстояния до 20 п.н. или уменьшение его до 15 п.н. приводит к тому, что РНК-полимераза не узнает испорченный промотор.
Этапы транскрипции
1. Узнавание и прочное связывание
Как только произошло узнавание (позиция 1), РНК-полимераза перемещается в позицию 2. В каталитическом центре инициации транскрипции, находящемся в -субъединице, оказывается +1-ый нуклеотид оперона. Переход из позиции 1 в позицию 2 возможен, если на операторе нет белка-репрессора.
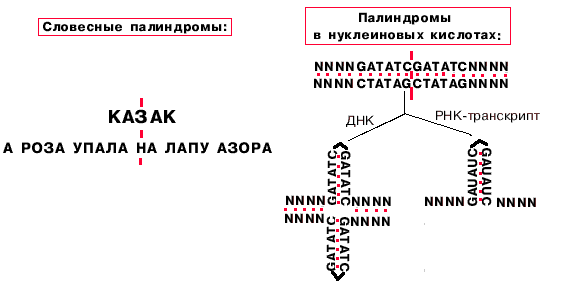
Примерно 5% промоторов у прокариот имеют только участок "-10", однако, тем не менее, хорошо узнаются РНК-полимеразой. Такие промоторы представлены палиндромными последовательностями, принимающими форму креста при суперспирализации кольцевых молекул ДНК.
Определение: палиндромы - последовательности, которые читаются одинаково слева направо и справа налево.
Палиндромы первого порядка имеют одну ось симметрии, второго - две, третьего - три.
2. Инициация заключается в образовании первой фосфодиэфирной связи между пурин-трифосфатом (АТФ или ГТФ) и следующим нуклеотидом. После инициации - фактор покидает фермент.
3. Элонгация - последовательное наращивание цепи РНК (или продолжение транскрипции).
Скорость элонгации 40-50 нукл./сек.
Для комплементарного синтеза РНК необходим разрыв водородных связей в ДНК. Core-фермент РНК- полимеразы покрывает примерно 40 пар нуклеотидов (4 витка спирали ДНК). Разрыв водородных связей на 4-х витках спирали - очень энергоемкий процесс. Он не был обнаружен при изучении транскрипции.
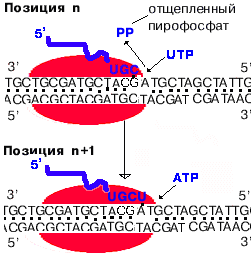 | Показано, что РНК-полимераза переводит ДНК из В-формы в А-форму. В ней плоскости азотистых оснований не перпендикулярны оси спирали, а наклонены на 200 к перпендикуляру. Это облегчает "выворачивание" двух соседних азотистых оснований в цепи ДНК для того, чтобы напротив них встали комплементарные нуклеотиды РНК. В пользу этого говорит полная идентичность параметров А-формы ДНК и гибрида, состоящего из одной цепи ДНК и одной - РНК. "Мотором" транскрипции является энергия, высвобождающаяся при отщеплении пирофосфата от каждого рибо-НТФ. |
Ингибиторы транскрипции прокариот.
Существует множество ингибиторов транскрипции. Они действуют по разным механизмам и на разных стадиях. Большинство из них - антибиотики.
Рифампицин - ингибитор инициации.
Связывается с центром инициации holo-РНК-полимеразы E. сoli.
Стрептолидигин - ингибитор элонгации.
Связывается с центром элонгации