Файл: Курс лекций, нгу 2000. Лекция Что такое молекулярная биология. Доказательства роли нуклеиновых кислот.doc
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 11.01.2024
Просмотров: 156
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
Определение: аминоацилирование - это образование связи между аминокислотой и tPHК.
Следующий этап трансляции - собственно синтез полипептидов, происходит на рибосомах.
Структура рибосом
Рибосомы - немембранные самые мелкие клеточные органеллы, при этом они едва ли не самые сложные. В клетке E. сoli присутствует около 103-5х103 рибосом. Линейные размеры прокариотической рибосомы 210 х 290 Å. У эукариот - 220 х 320 Å.
Выделяют четыре класса рибосом:
1. Прокариотические 70S.
2. Эукариотические 80S.
3. Рибосомы митохондрий (55S - у животных, 75S - у грибов).
4. Рибосомы хлоропластов (70S у высших растений).
Определение: S - коэффициент седиментации или константа Сведберга. Отражает скорость осаждения молекул или их компонентов при центрифугировании, зависящую от конформации и молекулярного веса.
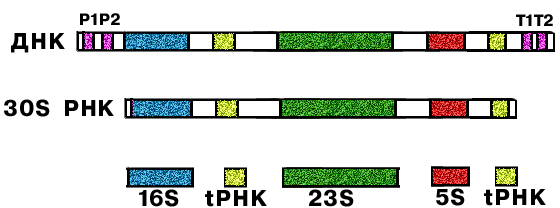
Каждая рибосома состоит из 2-х субъединиц (большой и малой).
| Прокариотическая рибосома | Эукариотическая рибосома | ||
| 70S | 80S | ||
| 50S | 30S | 60S | 40S |
| 5S rРНК 23S rРНК | 16S rРНК | 5S rРНК 5.8S rРНК 28S rРНК | 18S rРНК |
| 34 молекулы белков, из них 31 разные | 21 белок | не менее 50 разных белков | не менее 33 разных белков |
Сложность объясняется тем, что все элементы рибосом представлены в одном экземпляре, за исключением одного белка, присутствующего в 4 копиях в 50S субъединице, и не могут быть заменены.
rРНК выполняют не только функцию каркасов субъединиц рибосом, но и принимают непосредственное участие в синтезе полипептидов.
23S rРНК входит в каталитический пептидилтрансферазный центр, 16S rРНК необходима для установки на 30S субъединице инициирующего кодона mРНК, 5S rРНК - для правильной ориентации аминоацил-tРНК на рибосоме.
Все rРНК обладают развитой вторичной структурой: около 70% нуклеотидов собрано в шпильки.
rРНК в значительной степени метилированы (СН3-группа во втором положении рибозы, а также в азотистых основаниях).
Порядок сборки субъединиц из rРНК и белков строго определен. Субъединицы, не соединенные друг с другом, представляют собой диссоциированные рибосомы. Соединенные - ассоциированные рибосомы. Для ассоциации нужны не только конформационные изменения, но и ионы магния Mg2+ (до 2х103 ионов на рибосому). Магний нужен для компенсации отрицательного заряда rРНК. Все реакции матричного синтеза (репликация, транскрипция и трансляция) связаны с ионами магния Mg2+ (в меньшей степени - марганца Mn2+).
Каталитические центры рибосом
| Асп - центр специфического узнавания. Здесь происходит взаимодействие кодон-антикодон. Р-центр - пептидильный, донорный. Он является донором формилметионина при инициации, или пептидила при элонгации трансляции. А-центр - аминоацильный, акцепторный. Акцептирует формилметионин в самом начале или пептидил при элонгации трансляции. К-центр - каталитический (фермент пептидилтрансфераза). В К-центре задействована 23S rРНК и несколько белков большой субъединицы. |
Синтез полипептидов на рибосоме
У прокариот перед каждым геном и соответственно в mРНК перед копией каждого гена имеется лидерная последовательность.
| Она может быть разного размера (до 160 нукл.) и разной первичной структуры, но обязательно содержит полипуриновую последовательность Шайна-Дальгарно, которая комплементарна 3'-концевому участку 16S rРНК. Комплементарными могут быть 3-9 нуклеотидов. | 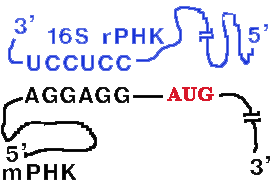 |
Назначение комплементарного взаимодействия 3'-концевого участка 16S rРНК и последовательности Шайна-Дальгарно - правильная установка инициирующего кодона AUG на малой субъединице рибосомы.
Инициирующий кодон находится на растоянии 3-10 нукл. от
последовательности Шайна-Дальгарно.
| К малой субъединице, на которой уже находится mРНК, подходит формилметиониновая tРНК, соединенная с формилметионином. В результате образуется инициаторный комплекс: 30S субъединица рибосомы + mРНК + формилметионовая tРНК-формилметионин. Затем происходит ассоциация рибосомы. При этом изменяется конформация 16S rРНК и нарушается связь между ней и последовательностью Шайна-Дальгарно. |  |
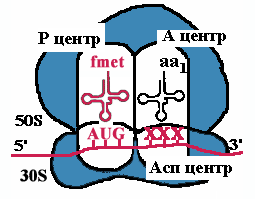 | Аминоацильный конец формилметиониновой tРНК оказывается в Р-центре. Второй кодон гена оказывается в Асп-центре. Соответствующая ему аминоацил-tРНК устанавливается таким образом, что ее аминоацильный конец попадает в А-центр. |
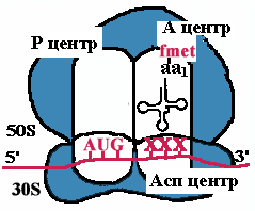 | Пептидилтрансфераза отрывает формилметионин в Р-центре и переносит его в А-центр. Образуется пептидная связь между формилметионином и аминоацил-tРНК. |
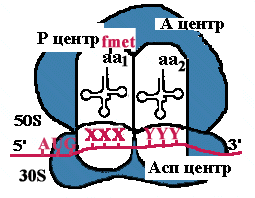 | Рибосома претерпевает конформационные изменения и сдвигается на один кодон. Формилметиониновая tРНК покидает рибосому. Второй кодон оказывается напротив Р-центра. Сюда же переходит tРНК, несущая на хвосте дипептид. В Асп-центр попадает третий кодон, а в А-центр очередная аминоацил-tРНК. |
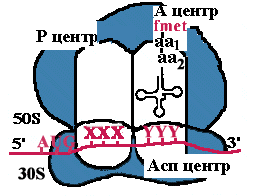 | Теперь в Р-центре отрывается дипептид, переносится в А-центр и соединяется с третьей аминоацил-tРНК. Так продолжается до тех пор, пока в Асп-центр не приходит терминирующий кодон. Полипептид отрывается в Р-центре, переносится в А-центр и, т.к. присоединиться ему не к чему, он отваливается от рибосомы. Рибосома диссоциирует и малая субъединица сканирует mРНК. |
In vivo на каждой стадии (образования инициаторного комплекса, инициации, элонгации и терминации) участвуют различные белковые факторы, которые препятствуют посадке на рибосому деацилированных tРНК или запрещают посадку формилметиониновой-tРНК в А-центр.
На всех этапах принимают участие молекулы ГТФ, которые дефосфорилируются.
Смысл гидролиза ГТФ не в отдаче энергии, а в свидетельстве того, что данный этап трансляции пройден.
| Все синтезируемые полипептиды прокариот на N-конце несут формилметионин. В 20% случаев он отщепляется, а в 80% отщепляется только формильная группа и на N конце остается метионин. | 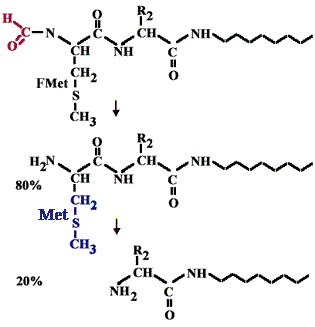 |
Регуляция образования рибосомных РНК и белков рибосом E.сoli
Ежеминутно в E.сoli образуется около 500 рибосом.
Имеется 7 оперонов, в которых закодированы rРНК (всего 3 разных rРНК х 7оперонов = 21 ген). В формировании рибосом участвуют 52 различных белка, а значит 52 гена, их кодирующих. В итоге,
73 гена должны работать координированно, чтобы не было избытка белков или rРНК.
Вначале образуется про-rРНК, которая метилируется и процессируется (т.е. "созревает").
 |
Количество rРНК регулируется количеством рибосомных оперонов, скоростью их транскрипции и работой ферментов метилаз и эндонуклеаз.
Имеется 7 разных оперонов, в которых закодированы рибосомные белки. Регуляция каждого из них осуществляется отдельно.
-оперон регулируется белком S4.
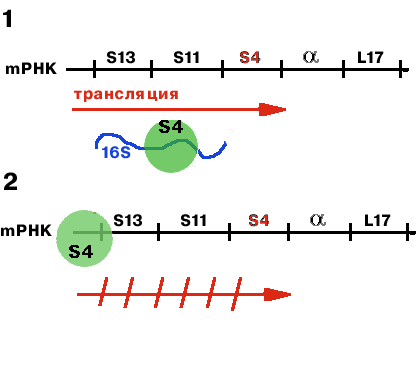 | Если в клетке имеется свободная 16S rРНК, то S4 связывется с ней (1). Если же 16S rРНК не хватает, то он связывается с mРНК, считывающейся с данного оперона (2). Причем связывается в районе лидера и тем самым мешает трансляции. |
Таким образом, осуществляется регуляция на уровне трансляции.
Оперон S10 регулируется белком L4.
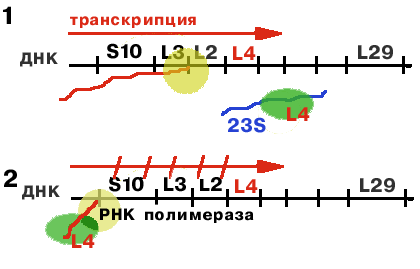 | РНК-полимераза синтезирует первую лидерную последовательность, длиной 140 нукл. Если 23S rРНК не хватает (2), то белку L4 не с чем соединяться, и он взаимодействует с лидерной последовательностью, придавая ей такую конформацию, которая не позволяет РНК-полимеразе продолжать транскрипцию. В результате синтез mРНК обрывается на первом же лидере (2). |
Регуляция на уровне транскрипции.
Оперон .
 |
В этом опероне закодированы белки, имеющие принципиальное значение для инициации транскрипции ( - фактор), инициации репликации (dna G - праймаза) и инициации трансляции (белок S21). Каждый белок нужен в разном количестве. S21 50000 копий, dnaG 50, - фактор 5000. Между геном S21 и геном dnaG есть слабый терминатор транскрипции. Ген dnaG имеет инициирующий кодон ГУГ (а не АУГ), который гораздо хуже узнается рибосомой и реже, чем АУГ инициирует трансляцию.
Аттенуация (ослабление)
Рассмотрим систему аттенуации на примере триптофанового оперона E.сoli.
Этот оперон регулируется по схеме негативной репрессии.
При недостатке в клетке триптофана оперон открыт. При увеличении концентрации триптофана РНК-полимераза не доходит даже до первого цистрона.
Между оператором и первым цистроном есть протяженный участок (162 п.н.), который содержит последовательность Шайна-Дальгарно. Она расположена ближе к цистрону, все остальное же представляет собой аттенуатор.
В этом районе происходит прекращение транскрипции и отсоединение РНК-полимеразы от ДНК. Это сделано для того, чтобы остановить РНК-полимеразу, которая уже в пути, в том случае, если концентрация триптофана в клетке к этому моменту повысилась.
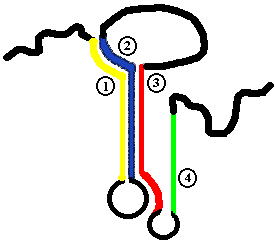 | В аттенуаторе выделяют 4 последовательности, частично комплементарные друг другу. В последовательности 1 закодирован 14-и членный пептид (Met-Lys-Ala-Ile-Phe-Val-Leu-Lys-Gly-Trp-Trp-Arg-Thr-Ser). На 10-ом и 11-ом месте в нем стоит триптофан. |