Файл: Курс лекций, нгу 2000. Лекция Что такое молекулярная биология. Доказательства роли нуклеиновых кислот.doc
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 11.01.2024
Просмотров: 159
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
| Если триптофан в клетке есть и доступен, то рибосома с легкостью преодолевает участок 1 и стерически мешает образованию шпильки (2)-(3). Тогда образуется шпилька (3)-(4), которая узнается РНК-полимеразой как сигнал прекращения транскрипции. Синтез mРНК обрывается. | 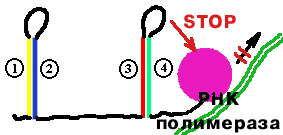 |
 | Если триптофан недоступен, то рибосома застревает на участке 1 и образуется шпилька (2)-(3). В этом случае не может образоваться шпилька (3)-(4). Сигнала для прекращения синтеза mРНК нет. |
Транскрипция у эукариот
У эукариот процессы транскрипции и трансляции разобщены во времени и пространстве (транскрипция - в ядре, трансляция - в цитоплазме).
У эукариот существуют специализированные РНК-полимеразы.
В ядре выделяют 3 типа РНК-полимераз:
РНК-полимераза I - синтезирует rРНК (кроме 5S rРНК).
РНК-полимераза II - синтезирует mРНК и некоторые sРНК.
РНК-полимераза III - синтезирует tРНК, некоторые sРНК и 5SrРНК.
РНК-полимеразы различаются количеством субъединиц, их аминокислотным составом, и зависимостью от катионов магния и марганца. Для РНК-полимераз I и III необходимое для работы соотношение [Mn2+]/[Mg2+] = 2. Для РНК-полимеразы II - [Mn2+]/[Mg2+] = 5.
Наиболее яркое различие - чувствительность к - аманитину (токсину бледной поганки). Он полностью подавляет работу РНК-полимеразы II в концентрации 10-8 М и РНК-полимеразы III ( в концентрации 10-6 М). РНК-полимераза I фактически нечувствительна к этому токсину.
Помимо ядерных РНК-полимераз у эукариот есть еще РНК-полимеразы хлоропластов и митохондрий. Они кодируются в ядре, а не в соответствующих органеллах.
В органеллах образуются свои tРНК, rРНК и рибосомные белки.
Как образуются рибосомы у эукариот
Гены rРНК присутствуют в количестве от 10 до 105 копий у разных видов (105
у амфибий). У человека - 300 генов, в которых закодированы rРНК.
Все рибосомные гены, кроме генов 5S рибосомной РНК, сближены (т.е располагаются один за другим) и образуют несколько кластеров. Сначала синтезируется про-rРНК, после созревания которой образуются 28S, 18S и 5,8S rРНК.
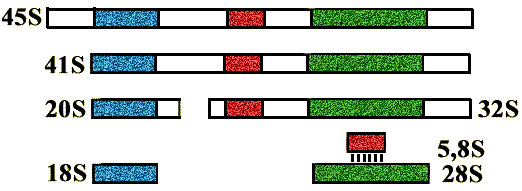
Интерфазные хромосомы в световой микроскоп не видны. Каждый ген прорибосомной РНК транскрибируется одновременно несколькими РНК-полимеразами и тут же начинается процессинг.
На электронномикроскопических фотографиях видна картина "рождественской елочки". Синтезируемые в ядре mРНК поступают на готовые рибосомы в цитоплазму, где синтезируются рибосомные белки, которые идут в ядро и путаются в "ветвях елки".
Образуются рибосомные субъединицы. Одновременно в эукариотическом ядре находятся сотни тысяч субъединиц рибосом.
Определение: ядрышко - место образования субъединиц рибосом, наблюдаемое в световой микроскоп.
В ядре может быть несколько ядрышек.
Определение: кластер генов rРНК называют ядрышковым организатором.
Особенности транскрипции эукариот
Единицей транскрипции у эукариот является отдельный ген, а не оперон, как у прокариот.
Оператор, как таковой, отсутствует.
Промотор есть, но он организован иначе.
На расстоянии -25 п.н. от +1 нукл. находится ТАТА-бокс. Его позиция определяет точку инициации транскрипции. А на расстоянии -60-80 п.н. находится ЦААТ-бокс, который не является абсолютно необходимым, но присутствует перед большинством генов.
Расстояние между ЦААТ и ТАТА большое и РНК-полимераза не способна накрыть всю эту область.
ЦААТ опознается своим белком, а ТАТА - своим.
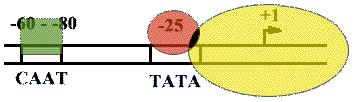
Помимо этих есть еще несколько белков, называемых базальными факторами транскрипции.
Определение: базальные факторы транскрипции - белки, необходимые для инициации транскрипции.
Базальные факторы транскрипции необходимы для инициации транскрипции всеми тремя ядерными РНК-полимеразами.
Для любого гена, кодирующего белок, есть энхансеры (усилители).
Определение: энхансеры - последовательности ДНК, усиливающие транскрипцию при взаимодействии со специфическими белками.
| Энхансеры - это не непрерывные последовательности нуклеотидов. Существуют так называемые модули - это отдельные части энхансеров. Одинаковые модули могут встречаться в разных энхансерах. Для каждого энхансера набор модулей уникален. Модули - это короткие последовательности, не более 2-х витков спирали (20 п.н.), которые могут находиться перед, за и даже внутри гена. | 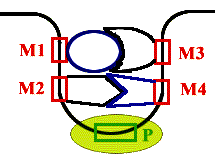 |
Таким образом, М1+М2+М3+М4 - один энхансер, но он состоит из 4-х модулей. Все 4 модуля узнаются своими белками, а они, сидя на ДНК, взаимодействуют друг с другом. Если в клетке присутствуют все соответствующие белки, то участку ДНК придается определенная конформация и начинается синтез mРНК.
Все соматические клетки многоклеточного эукариотического организма имеют абсолютно одинаковый набор генов. Почему же клетки дифференцированы и специализированы?
Дело в том, что все гены работают на фоновом уровне и не имеют фенотипического проявления. Экспрессируются лишь те гены, у которых все энхансерные модули узнаны своими белками и эти белки взаимодействуют друг с другом. Кроме энхансеров есть сайленсеры (ослабители).
Определение: сайленсеры - последовательности ДНК, ослабляющие транскрипцию при взаимодействии с белками.
При соответствующем наборе белков экспрессия отдельных генов в клетке может быть подавлена.
Процессинг (созревание) rРНК и tРНК у эукариот принципиально не отличается от такового у прокариот.
Процессинг mРНК отличается сильно и состоит из нескольких этапов.
1. Кепирование 100% mРНК
2. Полиаденилирование
95% mРНК
3. Сплайсинг
95% mРНК. Сплайсингу подвергаются только полиаденилированные mРНК.
4. Редактирование
Показано лишь для нескольких mРНК.
Все стадии процессинга mРНК происходят в РНП-частицах (рибонуклеопротеидных комплексах.
По мере синтеза про-mРНК, она тут же образует комплексы с ядерными белками - информоферами. И в ядерные, и в цитоплазматические комплексы mРНК с белками (информосомы) входят sРНК.
Таким образом, mРНК не бывает свободной от белков.
На всем пути следования до завершения трансляции mРНК защищена от нуклеаз. Кроме того, белки придают ей необходимую конформацию.
Определение: полисома - комплекс mРНК с несколькими или многими рибосомами.
В составе информосом mРНК может жить от нескольких минут до нескольких дней, не подвергаясь действию нуклеаз. (Так, mРНК живут неделями в ооцитах, предшественниках яйцеклеток).
Кепирование
Кепирование - надевание "шапочки".
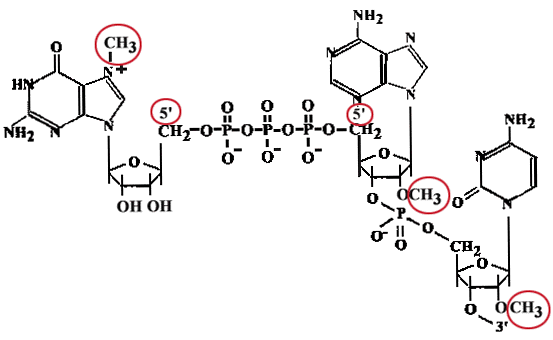
"Сар" представляет собой метилированный ГТФ, присоединенный в необычной позиции 5'-5' и две метилированные рибозы в первых двух нуклеотидах mРНК. По мере образования про-mРНК (еще до 30-ого нуклеотида), к 5'-концу, несущему пуринтрифосфат, присоединяется гуанин, после чего происходит метилирование.
ГТФ + гуанинтрансфераза (Е) Е
фГ + ф-ф
5'ф-ф-ф-Пур-ф-Х-ф-Y-ф-Z-...+E
фГ Г5'-ф-ф-ф-5'-Пур-ф-Х-ф-Y-ф-Z...
Таким образом, М1+М2+М3+М4 - один энхансер, но он состоит из 4-х модулей. Все 4 модуля узнаются своими белками, а они, сидя на ДНК, взаимодействуют друг с другом. Если в клетке присутствуют все соответствующие белки, то участку ДНК придается определенная конформация и начинается синтез mРНК.
Все соматические клетки многоклеточного эукариотического организма имеют абсолютно одинаковый набор генов. Почему же клетки дифференцированы и специализированы?
Дело в том, что все гены работают на фоновом уровне и не имеют фенотипического проявления. Экспрессируются лишь те гены, у которых все энхансерные модули узнаны своими белками и эти белки взаимодействуют друг с другом. Кроме энхансеров есть сайленсеры (ослабители).
Определение: сайленсеры - последовательности ДНК, ослабляющие транскрипцию при взаимодействии с белками.
При соответствующем наборе белков экспрессия отдельных генов в клетке может быть подавлена.
Процессинг (созревание) rРНК и tРНК у эукариот принципиально не отличается от такового у прокариот.
Процессинг mРНК отличается сильно и состоит из нескольких этапов.
1. Кепирование 100% mРНК
2. Полиаденилирование
95% mРНК
3. Сплайсинг
95% mРНК. Сплайсингу подвергаются только полиаденилированные mРНК.Таким образом, М1+М2+М3+М4 - один энхансер, но он состоит из 4-х модулей. Все 4 модуля узнаются своими белками, а они, сидя на ДНК, взаимодействуют друг с другом. Если в клетке присутствуют все соответствующие белки, то участку ДНК придается определенная конформация и начинается синтез mРНК.
Все соматические клетки многоклеточного эукариотического организма имеют абсолютно одинаковый набор генов. Почему же клетки дифференцированы и специализированы?
Дело в том, что все гены работают на фоновом уровне и не имеют фенотипического проявления. Экспрессируются лишь те гены, у которых все энхансерные модули узнаны своими белками и эти белки взаимодействуют друг с другом. Кроме энхансеров есть сайленсеры (ослабители).
Определение: сайленсеры - последовательности ДНК, ослабляющие транскрипцию при взаимодействии с белками.
При соответствующем наборе белков экспрессия отдельных генов в клетке может быть подавлена.
Процессинг (созревание) rРНК и tРНК у эукариот принципиально не отличается от такового у прокариот.
Процессинг mРНК отличается сильно и состоит из нескольких этапов.
1. Кепирование 100% mРНК
2. Полиаденилирование
4. Редактирование
Показано лишь для нескольких mРНК.
Все стадии процессинга mРНК происходят в РНП-частицах (рибонуклеопротеидных комплексах.
По мере синтеза про-mРНК, она тут же образует комплексы с ядерными белками - информоферами. И в ядерные, и в цитоплазматические комплексы mРНК с белками (информосомы) входят sРНК.
Таким образом, mРНК не бывает свободной от белков.
На всем пути следования до завершения трансляции mРНК защищена от нуклеаз. Кроме того, белки придают ей необходимую конформацию.
Определение: полисома - комплекс mРНК с несколькими или многими рибосомами.
В составе информосом mРНК может жить от нескольких минут до нескольких дней, не подвергаясь действию нуклеаз. (Так, mРНК живут неделями в ооцитах, предшественниках яйцеклеток).
Кепирование
Кепирование - надевание "шапочки".
"Сар" представляет собой метилированный ГТФ, присоединенный в необычной позиции 5'-5' и две метилированные рибозы в первых двух нуклеотидах mРНК. По мере образования про-mРНК (еще до 30-ого нуклеотида), к 5'-концу, несущему пуринтрифосфат, присоединяется гуанин, после чего происходит метилирование.
ГТФ + гуанинтрансфераза (Е) Е
(метилирование) Гm-ф-ф-ф-Пурm-ф-Хm-ф-Y-ф-Z-...
Назначение "Сар"
1. Защита 5'-конца mРНК от действия экзонуклеаз.
2. За счет узнавания "Сар"-связывающими белками происходит правильная установка mРНК на рибосоме.
Полиаденилирование
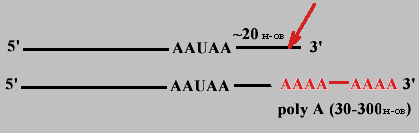
Когда синтез про-mРНК завершен, то на расстоянии примерно 20 нуклеотидов в направлении к 3' - концу от последовательности 5'-AAУAA-3' происходит разрезание специфической эндонуклеазой и к новому 3'-концу присоединяется от 30 до 300 остатков АМФ (безматричный синтез).
Каждый вид mРНК имеет "поли-А хвост" определенной длины. Он защищает 3'-конец от гидролиза, т.к. покрыт полиА-связывающими белками.
В значительной степени время жизни mРНК коррелирует с длиной полиА-хвоста.
mРНК ряда генов не полиаденилируется (например гистоновых генов).
Полиаденилированные про-mРНК подвергаются сплайсингу.
Сплайсинг
В 1978г. Филипп Шарп (Массачусетский технологический институт) открыл явление сплайсинга РНК (от англ. to splace - сшивать без узлов).
Определение: экзоны - кодирующие участки генов.
Определение: интроны - некодирующие участки генов.
На долю интронов приходится в 5-7 раз больше нуклеотидных пар, чем на долю экзонов. Количество экзонов в гене больше, чем интронов.
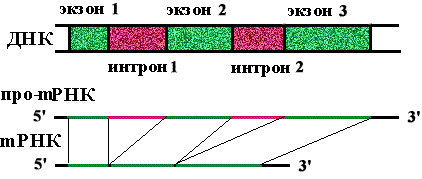
Определение: сплайсинг - вырезание копий интронов из про-mРНК и сшивание копий экзонов с образованием mРНК.
Копии интронов гидролизуются до нуклеотидов.
Сплайсинг показан для большинства mРНК и некоторых tРНК. У простейших найден автосплайсинг rРНК. Сплайсинг показан даже для археобактерий.
Не существует единого механизма сплайсинга. Описано по крайней мере 5 разных механизмов.
В ряде случаев сплайсинг осуществляют ферменты-матюразы.
В некоторых случаях в процессе сплайсинга участвуют sРНК.
В случае автосплайсинга процесс происходит благодаря третичной структуре про-РНК.
Для mРНК высших организмов существуют обязательные правила сплайсинга: