Файл: Воронова, Климова, Менджерицкий Анатомия центральной нервной системы.pdf
Добавлен: 19.02.2019
Просмотров: 6660
Скачиваний: 32

Рис. 28. Поперечный разрез продолговатого мозга.
Левая половина — схема расположения ядер (обозначены светлым) и проводящих путей (обозначены
темным):
1 — задний продольный пучок; 2 — шов; 3 — пирамида; 4 — корешок подъязычного нерва; 5 — медиальная
петля (осязательный путь); 6 — ядро оливы; 7 — ретикулярная формация; 8 — спиноталамический тракт; 9
— двойное ядро; 10— корешок блуждающего нерва; 11 — дорсальный продольный пучок; 12 — нижние
ножки мозжечка; 13 — ядро Дейтерса; 14 — одиночный (солитарный) тракт и его ядро; 15 — верхнее
вестибулярное ядро; 16 — дорсальное ядро блуждающего нерва; 17 — ядро подъязычного нерва
нимают промежуточное положение. Кроме ядер черепно-мозговых нервов в продолговатом мозге
расположены ядра, на которых переключается сенсорная информация (ядра олив, ядра Голля и
Бурдаха), и ядра ретикулярной формации, а также дыхательный, сосудодвигательный и
пищеварительные центры (рис. 28).
В продолговатом мозге и других стволовых отделах мозга (Варолиевом мосту и среднем мозге)
расположена особая клеточная структура — ретикулярная формация (рис. 28, 7). В
функциональном отношении ретикулярная формация ствола представляет собой единую
структуру. Название этой структуры отражает ранние представления о том, что отдельные
нейроны ретикулярной формации имеют обширные связи друг с другом и образуют нечто
подобное нейропилю, в котором возбуждение распространяется диффузно,
55
подобно тому, как это происходит в нервной системе кишечнополостных. Однако в дальнейшем
эти представления не подтвердились.
Четких границ между отдельными ретикулярными и неретикулярными группами клеток не
установлено, однако выделено до 98 ядерных групп, относящихся к ретикулярной формации.
Основными ядрами являются: ядра шва и гигантоклеточное ядро продолговатого мозга,
центральное и ретикулярное ядра моста.
Клетки ретикулярной формации очень разнообразны по форме и величине. Для них характерно
наличие значительно разветвленного дендритного дерева и длинных аксонов. Афферентные входы
ретикулярная формация получает как по коллатералям восходящих (сенсорных) путей, так и от
вышележащих структур, в том числе от коры больших полушарий и мозжечка. Таким образом,
ретикулярная формация интегрирует влияние большого числа мозговых структур. В свою очередь
сама она оказывает влияние как на вышележащие, так и на нижележащие структуры.
Нисходящие и восходящие волокна ретикулярной формации покидают ядра на всем ее
протяжении, не имея четких пространственных разграничений. Кроме того, существуют аксоны,
которые Т-образно ветвятся. Одна из ветвей идет вниз, а другая — в верхние отделы головного
мозга.
Нисходящие волокна образуют ретикулоспинальный тракт. Через него ретикулярная формация
оказывает влияние как на двигательную деятельность спинного мозга (осуществление спинальных
рефлексов), так и на вегетативную регуляцию (сосудодвигательная, дыхательная,
пищеварительная функции). Ретикулярная формация воздействует на соматические и
вегетативные центры в двух противоположных направлениях: торможения и возбуждения.
Восходящие влияния ретикулярной формации направлены на регуляцию деятельности коры

больших полушарий. Большинство ретикулярных волокон достигает коры через переключение в
неспецифических ядрах таламуса. Действие восходящих ретикулярных влияний заключается в
широкой активации корковых структур. Торможение активности ретикулярной формации ведет к
наступлению сна, активация ее приводит к реакции пробуждения.
Характерной особенностью нейронов ретикулярной формации является их высокая
чувствительность к химическим факторам. Такие факторы, как уровень углекислого газа и
кислорода, содержание адреналина, ацетилхолина и серотонина, относительно небольшие
концентрации фармакологических веществ, изменяют актив-
56
ность нейронов ретикулярной формации, а вместе с этим и ее влияние на кору больших
полушарий, соматические и вегетативные рефлексы.
Филогенез.
Филогенез. В филогенезе развитие продолговатого мозга определяется развитием чувствительных
органов боковой линии, слуха, тактильной чувствительности тела. У всех позвоночных животных,
кроме млекопитающих, в продолговатом мозге расположены ядра V— XII пар нервов. Развитие
органов боковой линии и слуха определяет утолщение дорсальной части боковой стенки
продолговатого мозга. У человека из этих структур остаются слуховые бугорки.
Вентральные структуры продолговатого мозга развиваются благодаря связям вышележащих
отделов (коры больших полушарий) с двигательными структурами спинного мозга. Поэтому у
низших позвоночных пирамиды отсутствуют, а у млекопитающих становятся ясно различимы.
Оливы появляются только у высших обезьян. Поскольку наибольший кортикальный контроль над
двигательными структурами наблюдается у человека, у него же наиболее выражены и пирамиды,
и оливы.
Филогенетически ретикулярная формация является древней структурой. У низших позвоночных
она распространена по всей центральной нервной системе. У высших позвоночных эта структура
хорошо представлена только в стволовых отделах мозга, так как функцию регуляции берут на себя
конечный мозг и мозжечок.
Онтогенез.
Онтогенез. В онтогенезе продолговатый мозг является производным ромбовидного мозгового
пузыря (rhombencephalon), который делится на задний (metencephalon) и добавочный мозг
(myelencephalon). Полость ромбовидного мозга не делится и остается общей для продолговатого и
заднего мозга — ГУ желудочком. Добавочный мозг развивается в продолговатый. Продолговатый
мозг формируется так же, как и спинной. С дорсальной стороны он имеет крыловидные
пластинки, между ними крышу, с вентральной стороны — базальные пластины и дно. Однако
крыловидные пластинки расходятся, крыша растягивается и становится очень тонкой. В ней
образуется сосудистое сплетение, которое заметно уже на 6-й неделе развития.
В структурах продолговатого мозга формируются ядра черепно-мозговых нервов. Причем почти
все ядра нервов (тройничного, отводящего, лицевого, слухового), принадлежащие во взрослом
состоянии мосту, закладываются первоначально в ростральных отделах продолговатого мозга, и
только позже они перемещаются в структуры заднего мозга.
57
Сосудистое сплетение IV желудочка на начальных этапах эмбрионального развития человека
представляет собой сплошную пластинку. В дальнейшем она прорывается, образуя отверстия:
места сообщения между полостью IV желудочка и полостью подпаутинного пространства.
6.2.2. Задний мозг
Задний мозг развивается из заднемозгового пузыря (теtencéphalon), являющегося производным
ромбовидного пузыря (rhombencephalon). Вентральная часть заднего мозга представляет собой
продолжение стволовых структур и называется Варолиев мост. Варолиев мост несет в своем
составе полость заднего мозга — часть ромбовидной ямки. Крыша ромбовидной ямки
претерпевает значительные изменения и развивается в мозжечок, представляющий собой
дорсальный вырост заднего мозга.
6.2.2.1. Варолиев мост
Варолиев мост (pons Varolii) — это вентральная часть заднего мозга. Сам мост образует структуры
ростральной части дна IV желудочка. Дорсальная поверхность моста представляет собой верхний

треугольник ромбовидной ямки. Полость ромбовидной ямки рострально сужается и переходит в
водопровод среднего мозга. Сверху полость ромбовидной ямки прикрыта верхним мозговым
парусом, который вместе с нижним мозговым парусом и сосудистым сплетением образует крышу
IV желудочка, имеющую вид шатра. Латеральные стенки IV желудочка в области моста
образованы средними и верхними ножками мозжечка (см. рис. 25, 33).
Вентральная поверхность Варолиева моста представляет собой мощную поперечно-волокнистую
выпуклость белого вещества. По центру вентральной поверхности моста проходит глубокая
борозда — канавка основной артерии мозга (suclus basillaris). Латерально вентральная выпуклость
переходит в мощные средние ножки мозжечка (см. рис. 25, 5; 27, 4).
От Варолиевого моста отходят четыре пары черепно-мозговых нервов (см. рис. 26; 27).
V — тройничный нерв (п. trigeminus);
VI — отводящий нерв (п. abducens);
VII — лицевой нерв (п. facialis);
VIII — предверно-улитковый, или слуховой, нерв (п. vestibulocochlearis).
58
На поперечных срезах, как и в продолговатом мозге, видны белое вещество и ядра серого
вещества. Поперечные волокна, составляющие трапециевидное тело, делят толщу моста на более
крупную вентральную (основание моста) и дорсальную (покрышка моста) части. В вентральной
части преобладает белое вещество проводящих путей, являющихся продолжением проводящих
путей ножек среднего мозга. Серое вещество вентральной части моста образует собственные ядра
моста (ядра основания моста). В этих ядрах заканчиваются идущие из коры больших полушарий
нисходящие кортикомостовые пути и коллатерали от кортикоспинальных путей. От собственных
ядер моста отходят волокна, которые переходят на противоположную сторону и образуют
трапецевидные тела, переходящие в средние ножки мозжечка.
Дорсальная часть моста является непосредственным продолжением продолговатого мозга. В ней
располагаются переключательные ядра сенсорных систем, ядра черепно-мозговых нервов и
ретикулярной формации.
В филогенезе
В филогенезе у низших позвоночных животных Варолиев мост не отделен четко от
продолговатого мозга. Он обособляется только у млекопитающих. Это происходит с развитием
коры и нисходящих из нее проекционных путей. Вместе с этим растет количество собственных
ядер вентральной части моста. Это обусловливает появление и развитие средних ножек мозжечка
и его полушарий. Особенно значительно вентральные части моста и средние ножки мозжечка
выражены у человека.
В онтогенезе
В онтогенезе мост, как отдел заднего мозга, происходит из ромбовидного мозгового пузыря. На
стадии пяти мозговых пузырей ромбовидный мозг делится на добавочный (myelencephalon), из
которого развивается продолговатый мозг, и задний мозг (теtencéphalon). Крыша заднего мозга
преобразуется в мозжечок, а дно и стенки его становятся структурами моста. Полость
ромбовидного мозга остается общей для продолговатого мозга и моста и является полостью IV
желудочка.
Почти все ядра черепно-мозговых нервов моста закладываются в ростральных отделах
продолговатого мозга. Их перемещение в мост происходит после образования мозгового изгиба.
На 7-й неделе развития эмбриона клетки крыловидной пластинки продолговатого мозга
мигрируют в ростровентральном направлении и образуют на вентральной поверхности моста
понтобульбарное тело, которое позже превращается в собственные ядра моста.
59

6.2.2.2. Мозжечок
Рис. 29. Расположение мозжечка на стволе мозга (часть ткани мозжечка убрана):
1 — верхняя ножка мозжечка; 2 — средняя ножка мозжечка; 3 — лист (долька мозжечка); 4 — левое
полушарие мозжечка; 5 — спинной мозг; 6 — продолговатый мозг; 7 — нижняя ножка мозжечка; 8 — мост; 9
— средний мозг
Мозжечок (cerebellum) располагается на дорсальной поверхности ствола мозга. Вентральная
поверхность мозжечка прилежит к парусам IV желудочка и тесно связана со стволовыми
структурами тремя парами мозжечковых ножек: с продолговатым мозгом нижними ножками
(pedunculus cerebellaris inferior) (рис. 29, 7), с мостом средними ножками (pedunculus cerebellaris
médius) (рис. 29, 2) и со средним мозгом — верхними ножками (pedunculus cerebellaris superior)
(рис. 29, 1). Наиболее мощными являются средние ножки. Все ножки выходят из мозжечка рядом,
а потом верхние ножки направляются к среднему мозгу вместе с верхним мозговым парусом, а
нижние — к продолговатому мозгу вместе с нижним мозговым парусом. Рострально над
мозжечком лежат затылочные доли большого мозга, которые заходят за его дорсальные границы
мозжечка. Мозжечок отделен от большого мозга глубокой поперечной щелью мозга. Он, как и
большой мозг, покрыт тремя оболочками.
Анатомически мозжечок человека состоит из трех основных частей: двух полушарий (hemi-spheria
cerebelli) и соединяющей их средней части — червя (vermis cerebélli). Поверхность мозжечка
изрезана глубокими ветвящимися бороздами. Глубокие борозды мозжечка делят полушария и
червь на дольки, которые объединяются в доли: верхние, задние и нижние. Доли отделяются друг
от друга щелями.
Между обоими полушариями мозжечка располагается изрезанная узкими, параллельно идущими
бороздами средняя часть — червь (см. рис. 30а, 13). На нем различают верхнюю поверхность —
верхний червячок, и нижнюю — нижний червячок. Две продольно идущие бороздки на каждой
поверхности мозжечка отделяют верхний и нижний червячки от полушарий
60
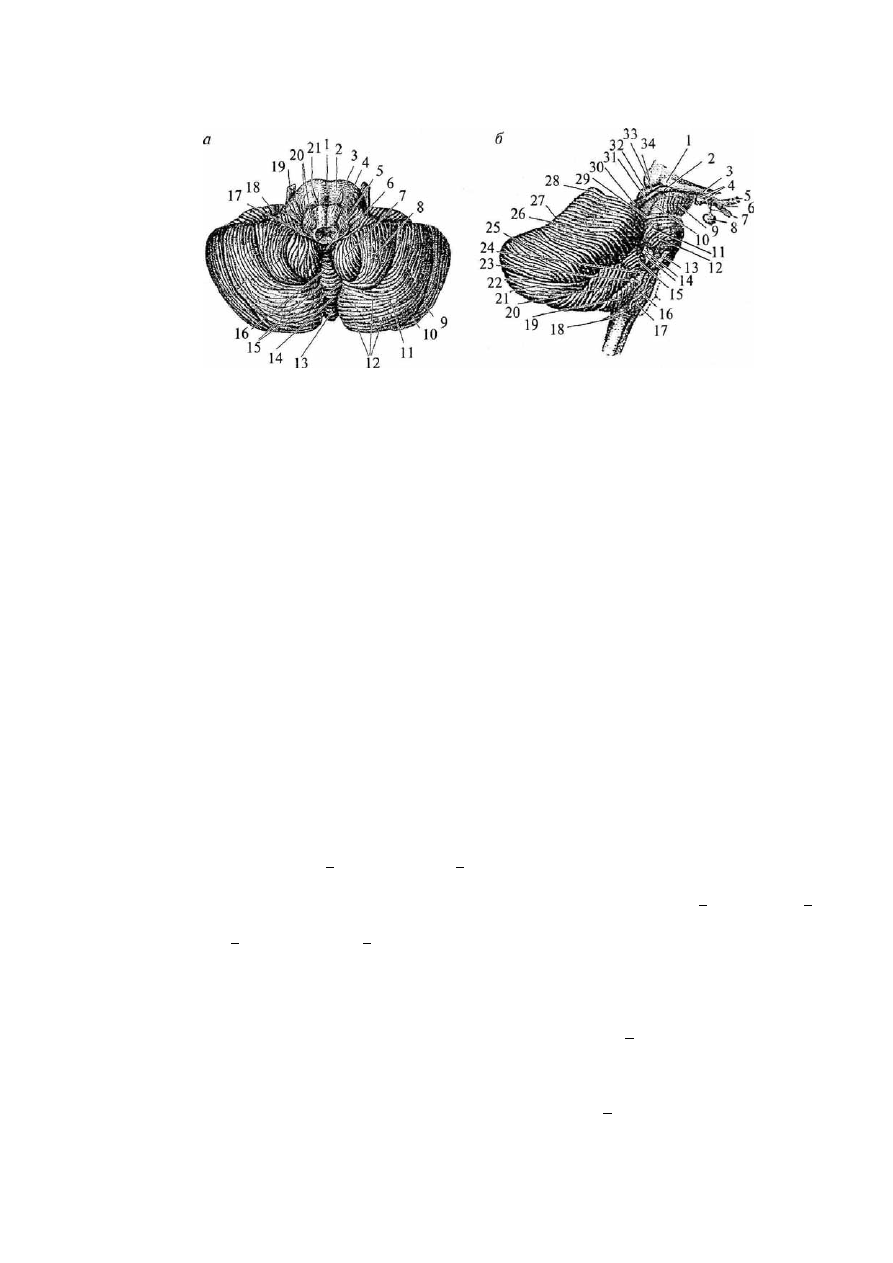
Рис. 30. Мозжечок:
вид снизу (о): 1 — борозда основной артерии; 2 — Варолиев мост; 3 — пирамида продолговатого мозга; 4 —
олива; 5 — узелок червя; 6 — сосудистое сплетение IV желудочка; 7 — миндалина мозжечка; 8 —
двубрюшная долька мозжечка; 9 — верхняя полулунная долька мозжечка; 10 — горизонтальная борозда
мозжечка; 11 — нижняя полулунная долька; 12 — извилины мозжечка; 13 — бугор червя; 14 — пирамида
червя; 15 — долинка мозжечка; 16 — язычок червя; 17 — ножка клочка; 18 — клочок; 19 — тройничный
нерв; 20 — корешки языкоглоточного и блуждающего нервов; 21 — отводящий нерв;
вид справа (б): 1 — латеральное коленчатое тело; 2 — медиальное коленчатое тело; 3 — зрительный тракт;
4 — мамиллярные тела; 5 — зрительный нерв; 6 — хиазма; 7 — воронка; 8 — гипофиз; 9 — ножка мозга; 10
— латеральная борозда среднего мозга; 11 — тройничный нерв; 12 — косой пучок моста; 13 — отводящий
нерв; 14 — слуховой и лицевой нервы; 15 — клочок; 16 — олива; 17 — подъязычный нерв; 18 — наружные
дугообразные волокна; 19 — миндалина мозжечка; 20 — двубрюшная долька мозжечка; 21 — нижняя
полулунная долька мозжечка; 22 — горизонтальная борозда мозжечка; 23 — блуждающий нерв; 24 —
языкоглоточный нерв; 25 — верхняя полулунная долька; 26 — четырехугольная долька; 27 — скат; 28 —
вершина; 29 — борозда блокового нерва; 30 — нижние бугры четверохолмия; 31 — нижние ручки
четверохолмия; 32 — верхние бугры четверохолмия; 33 .— верхние ручки четверохолмия; 34 — подушка
мозжечка. На верхнем и нижнем червячках различают доли, которые состоят из нескольких
извилин (рис. 29, 3).
Верхний червячок спереди назад состоит из следующих долей:
1) язычок мозжечка (рис. 30а, 16);
2) центральная долька (рис. 33, 23);
3) бугор (рис. 30а, 13);
4) лист червя, в виде очень узкой дольки располагающейся сзади, на границе перехода верхнего
червячка в нижний (рис. 29, 3).
61
На нижнем червячке, в направлении спереди назад, различают следующие доли:
1) узелок (рис. 33, 20, к передним отделам которого присоединяется задний мозговой парус;
2) втулочка червя;
3) пирамида червя (рис. 33, 16).
На верхней поверхности полушарий мозжечка отмечают следующие борозды и дольки.
Четырехугольная долька (lobulus quadrangularis) (рис. 30б, 26) разделяется передней верхней
бороздой (sulcus superior anterior) на переднюю и заднюю части. Четырехугольная долька
ограничивается задней верхней бороздой от верхней полулунной дольки (lobulus semilunaris
superior) (рис. 306, 25).
Впереди от lobulus quadrangularis залегают небольшие извилины, так называемые крылья
центральной дольки. Снизу и спереди от последних лежат небольшие участки мозжечка — связи
язычка (vinculo lingulae).
На нижней поверхности полушарий мозжечка располагаются следующие борозды и дольки.
Группа концентрически расположенных извилин образует миндалину (tonsilla) (рис. 30а, 7; 30б,
19). Снаружи и сзади от миндалины залегает двубрюшная долька (lobulus bivénter) (рис. 30а, 8,
30б, 20). Двубрюшная долька соответствует пирамиде червячка.
Снаружи от миндалины и спереди от двубрюшной дольки располагается наиболее выступающая
спереди часть нижней поверхности мозжечка — клочок (flocculus) (рис. 30а, 18; 306, 15).
Сзади от двубрюшной дольки лежит нижняя полулунная долька (lobulus semilunaris inferior) (рис.
30а, 11; 30б, 21).
Нейронная организация мозжечка существенно отличается от таковой стволовых структур.
Основная масса нейронов сосредоточена на поверхности и создает кору мозжечка (cortex