ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 31.10.2019
Просмотров: 807
Скачиваний: 1

ЛФ ФИУ ПФ Занятие № 1116
За открытие феномена «двойного распознавания» (1974 год) Цинкернагель и Дохерти получили в 1996 году
Нобелевскую премию.
Т-клеточный рецептор – это гетерополимер, состоящий из двух полипептидных цепей,
и
, имеющих оди-
наковую длину. Каждая цепь имеет короткий цитоплазматический участок, необходимый для проведения
сигнала активации внутрь лимфоцита, а также трансмембранный участок, состоящий из гидрофобных ами-
нокислот. TCR тесно связан с группой белков, называемой молекулой СD3. СD3 состоит из одной
, одной
, двух
и двух
ξ
цепей. Комплекс
+
+
+
составляет мембранную структуру
CD3.
Таким образом, в целом комплекс Т-клеточный рецептор – молекула СD3 состоит из 8 полипептидных це-
пей:
–
2 локализованы на мембране –это Ag-связывающая часть Т-клеточного рецептора
(она вовлечена в двойное распознавание антигена)
–
4 полипептидных цепи, также локализованных на мембране, принадлежат молекулам CD3 (их функ-
ция – правильная конформация и экспрессия Ag-связывающей части)
–
2 цепи находятся в цитоплазме, их роль – проведение сигнала внутрь Т-лимфоцита
В процессе участвуют также Т-клеточныекорецепторные молекулы:
–
CD8 – для взаимодействия c молекулами MHC-I
–
CD4 – для взаимодействия cмолекулами MHC-II
Рис. 29.2-1. КомплексТ-клеточный рецептор – молекула СD3.

ЛФ ФИУ ПФ Занятие № 1117
Рис. 29.2-2. Корецепторные молекулы Т-лимфоцитов – молекулы СD4 иСD8 .
29.3. Экспрессия поверхностных маркеров Т-лимфоцитов в процессе лимфопоэза и селекция ненужных кло-
нов
Лимфоциты —
это
клетки с двойной дифференцировкой (созреванием):
• первый этап происходит в центральных органах иммунной системы и не зависит от антигенного раздра-
жения. Этот процесс называют
лимфопоэзом.
Он заканчивается образованием основных субпопуляций Т-
лимфоцитов и формированием на их поверхности антигенраспознающих рецепторов;
• вторичная дифференцировка идет в периферических органах иммунной системы. Она индуцируется
антигеном, следовательно, антигензависима. Ее итогом является образование функционально различных
клеток.
Предшественники Т-лимфоцитов, несут на своей поверхности молекулы СD2. Поступая в тимус, под влия-
нием гормонов этого центрального лимфоидного органа они приобретают новые молекулы – кластеры диф-
ференцировки – СD3, СD4 и СD8. На данном этапе еще незрелые Т-лимфоциты (тимоциты) называют
двойными позитивными клетками, т.к. на их поверхности экспрессированы не только СD2 и СD3, но и сра-
зу два вида кластеров СD4 и СD8, а затем появляется еще и Т-клеточный рецептор. В клетках тимуса такие
двойные позитивные тимоциты проходят отбор путем связывания с молекулами МНС, несущими свой пеп-
тидный антиген, и локализованными на поверхности клеток тимуса. Тимоциты, которые не связываются с
МНС, погибают в результате апоптоза, как не способные вступать во взаимодействия с этим комлексом.
Данный процесс называют позитивной селекцией тимоцитов. Тимоциты, которые, напротив, связываются с
молекулами МНС с высокойаффинностью, также уничтожаются, поскольку они слишком активно реагиру-
ют с собственными антигенами МНС и могут дать начало аутореактивным клонам. В данном случае проис-
ходит негативная селекция тимоцитов. И, наконец, те тимоциты, которые связываются со среднейаффинно-
стью (не слишком активно) с молекулами МНС клеток тимуса, выживают и далее проходят через отбор на
способность связываться посредством Т-клеточного рецептора с собственными молекулами МНС I, либо II
класса, локализованными на эпителиоидных клетках тимуса. Те клетки, которые связываются при помощи
молекул СD4 с МНС II макрофагов тимуса, теряют молекулы СD8, и наоборот незрелые лимфоциты, пред-
почитающие связываться посредством взаимодействия их молекул СD8 с МНС I эпителиальных клеток ти-
муса, теряют СD4. В результате отбора возникает две субпопуляции уже зрелых, но еще неиммунных Т-
лимфоцитов: цитотоксические лимфоциты, несущие только молекулы СD8, и имеющие сродство к МНС I,
и Т хелперы, сохранившие только СD4-молекулы и имеющие сродство к МНС II.

ЛФ ФИУ ПФ Занятие № 1118
Рис.29.3-1. Лимфопоэз и селекция клонов лимфоцитов в тимусе.
Таким образом, в процессе лимфопоэза в тимусе формируются несколько субпопуляций Т-лимфоцитов, ко-
торые отличаются друг от друга маркерами CD, и которые впоследствии будут выполнять разные функции,
взаимодействуя с разными антигенами.
Следует помнить, что все Т-лимфоциты вне зависимости от того, к какой популяции они
принадлежат, несут на своей мембране молекулы
CD3
. Клоны лимфоцитов, имеющие кластеры
дифференцировки
CD4
называют
Т-хелперы
и обозначают как
Th
,
причем существуют
субпопуляции хелперов:
Th
1
,Th
2
и
Th
3
. В противоположность им те клоны Т-лимфоцитов,
которые экспрессируют на мембране молекулы
CD8
, называют
Т-киллеры/супрессоры.
Их
делят на субпопуляции по факту наличия молекул
CD28
(по North M.E., 1998):Т-киллеры –
CD8
+
CD28
+
иТ-супрессоры –
CD8
+
CD28
-
.
29.4. Следствия контакта с МНС зрелых Т-лимфоцитов
Зрелые, но неиммунные Т-лимфоциты поступают в периферические лимфоидные органы.Здесь в Т-зависимых
зонах они вступают с помощью Т-клеточного рецептора в контакт с чужеродным и собственным антигенми,
представленными в виде пептида в составе с комплексом МНС. Те лимфоциты, которые «узнают» чужерод-
ный антиген получают сигнал к пролиферации, активации, синтезу и секреции биологически активных ве-
ществ – цитокинов, индуцируя таким образом развитие иммунологических реакций, направленных на защиту
организма от чужеродного антигена. Лимфоциты, которые распознают собственные антигены организма, так-
же представленные на поверхности дендритных клеток лимфоидных органов в составе с комплексом МНС,
получают «сигнал на выживание» и начинают пролиферацию. Те лимфоциты, которые не способны вступать
в контакт с комплексом МНС – антиген, погибают, поскольку они не способны воспринимать основной сиг-
нал, активирующий развитие иммуного ответа.

ЛФ ФИУ ПФ Занятие № 1119
Рис.29.4-3. Селекция зрелых Т-лимфоцитов в периферических лимфоидных органах.
29.5. Функции Т-лимфоцитов
Т-лимфоциты
в процессе дифференцировки и пролиферации образуют субпопуляции, отличающиеся друг
от друга по своим функциям: одни выполняют
регуляторные,
а другие —
эффекторные функции.
К регуля-
торам относят
Т-хелперы (Th)
(от англ. helper) илипомощники. Среди них различают следующиесубпопу-
ляции:
Th1
выделяют цитокины (ИЛ-2, ИЛ-12,
-интерферон) под действием которыхактивируется образование эф-
фекторных клеток —
Т-киллеров
, лимфоцитов, осуществляющих реакции клеточного иммунитета и стиму-
лирующих иммунитет
против вирусов и внутриклеточных бактерий
.
Th2
выделяют ИЛ-4, ИЛ-5, ИЛ-6 и стимулируют В-лимфоциты, способствуя трансформации В-лимфоцитов
в плазматические клетки, клетки – продуценты антител (активируют реакции гуморального иммунитета).
Антитела участвуют в защите организма от
бактерий, бактериальных токсинов, вирусов
и других пато-
генов.
Тh3
относятся к регуляторным клеткам: они регулирует функции Th1 и Th2, выделяют ИЛ-10 (супрессорный
фактор), TGF-
(трансформирующий фактор роста-
), регулируя развитие как клеточного, так и гумораль-
ного иммунного ответа.
Известны также:
T-reg
(от англ. Т-regulator) или
Т-регуляторные клетки
–клетки
CD4
+
CD25
+
. Онисодержат
белок Foxp3
,
который подавляет активность других Т-клеток;
Тs
(от англ.T-supressor) или
T-супрессоры
тормозят слишком сильные и слишком затянувшиеся иммунологи-
ческие реакции.
К эффекторным Т-лимфоцитам относят:
Tk
(от англ. Т-killer) или
Т-киллеры (убийцы)
– лимфоциты, которые уничтожают злокачественные, переса-
женные, мутантные, модифицированные, стареющие, пораженные вирусами и бактериями клетки.
Т-эффекторы воспаления
– осуществляют клеточные иммунные реакции воспаления.
Из общей популяции Т-лимфоцитов выделяют также
Т-клетки иммунологической памяти
– это долгожи-
вущие Th и Tk, потомки клеток, встречавшихся с Ag и сохранивших к ним рецепторы. Эти долгоживущие
антигенстимулированныеклетки обеспечивают более быстрый и выраженный иммунный ответ при повтор-
ной встрече с тем же антигеном —
вторичный иммунный ответ.
29.6. BCR – рецептор В-лимфоцита
Рецептор В-лимфоцита
-
BCR (
от англ. B – cellularreceptor
)
способен распознавать непроцессированный ан-
тиген. B-лимфоциты используют в качестве рецептора связанный на поверхности клетки иммуноглобулин.
Специфичностьэтогорецепторатакаяже, какспецифичностьиммуноглобулина,
которыйлимфоцитысекретируютпослеихактивации. В-
лимфоцитыспособныраспознаватьследующиеантигены, представленныеврастворимойформе:
1)
белки (какконформационныедетерминанты, такидетерминанты, образовавшиеся после
денатурации)
2)
нуклеиновыекислоты
3)
полисахариды
4)
некоторыелипиды
5)
небольшие по молекулярной массе соединения (гаптены)
ЛФ ФИУ ПФ Занятие № 1120
Зрелые В-лимфоциты, которые проходят все стадии созревания в костном мозге, экспрессируют на мембра-
не так называемые мембранные иммуноглобулины:
IgM
и
IgD
.
Первым клеточным рецептором, появляю-
щимся на В-лимфоците, является IgM. В отличие от секретируемого плазматической клеткой в виде анти-
тел пентамерногоIgM рецепторный иммуноглобулин является мономером, поэтому его часто обозначают
как mIgM.Он всегда обнаруживается на мембране В-лимфоцита. Главная функция В-клеточного рецептора
– связывание с антигеном и проведение сигнала активации в клетку В-лимфоцита. Для этого у рецепторного
IgM имеется короткий цитоплазматический хвост, вовлеченный во взаимодействие с комплексом, который
осуществляет проведение сигнала активации (сигнализирует о появлении антигена и необходимости им-
мунного реагирования на него) в клетку. Комплекс представлен четырьмя трансмембранными полипепти-
дами (рис.29.6-1).
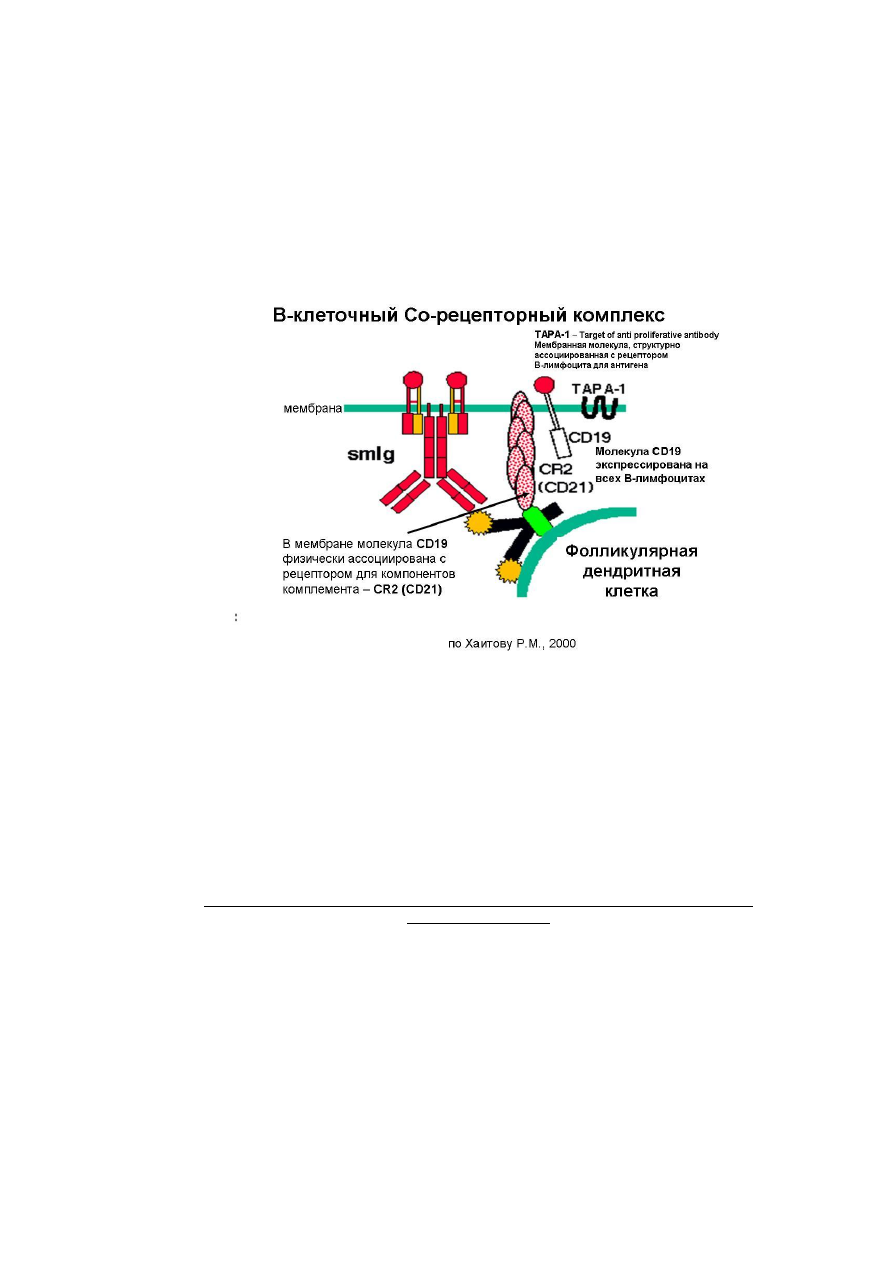
Рис.29.6-1. В-клеточный Со-рецепторный комплекс (по Хаитову Р.М., 2000
).
Для эффективного функционирования В-лимфоцитов существенна экспрессия на мембране
корецепторных молекул
CD19
и
CD21
. Молекула
CD19
экспрессирована на всех В-лимфоцитах.
В мембране молекула
CD19
физически ассоциирована с
CR2 (CD21
) – рецептором для
связывания компонентов комплемента. Встроенная в мембрану В-лимфоцита
TAPA-1
(от англ.
targetofantiproliferativeantibody) является мембранной молекулой, структурно ассоциированной с
рецепторомВ-лимфоцита дляантигена. Мембранные молекулы
CD19
,
CD21
и
CR2 (CD21
)
в диагностике используются как маркеры для определения В-лимфоцитов лабораторными
методами.
29.7. Экспрессия BCR в процессе лимфопоэза и негативная селекция аутореактивных
клонов В-лимфоцитов
А.
Созревание В-лимфоцитов
.
Процесс созревания происходит в костном мозге. Здесь идет поэтапное появление на мембране лимфоцита
иммуноглобулиновых рецепторов. ПервымэкспрессируетсяIgM и формируется незрелый В-лимфоцит. В
случае контакта на этой стадии с Ag происходят следующие события:
апоптоз («делеция клона») – гибель В-лимфоцитов
анергия (блок проведения сигнала с мембраны внутрь В-лимфоцитов)
изменение специфичности BCR(«редакция» рецептора по антигенной специфичности).
На следующем этапеидет формированиезрелого В-лимфоцита, на мембране которогоэкспрессируетсяIgD. В
случае контакта с антигеном на этой стадии происходит:
активация В-лимфоцита
его пролиферация