ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 27.11.2019
Просмотров: 4031
Скачиваний: 98

Элиситориндуцируемый
синтез ферментов, катализиру-
ющих образование стрессовых
фитогормонов. Установле-
но, что патогены и элиситоры вызывают экспрессию генов
ферментов 1 -аминоциклопропан-1 -карбоксилат-синтазы
[Liu et al.,1998]
и 1-аминоциклопропан-1-карбоксил ат-окси-
дазы [Jia, Martin, 1999], катализирующих реакции образова-
ния этилена. Повышение концентрации этилена активиро-
вало 1 -аминоциклопропан-1-карбоксилат-синтазу [J. Arteca,
R. Arteca,
1999], что может быть еще одним примером
отмечавшихся нами ранее [Гречкин, Тарчевский, 1999; Тар-
чевский, 2000] явлений автокатализа сигнального метабо-
лизма.
Элиситориндуцируемый синтез жасмоната происходит в
результате активации экспрессии генов ферментов липок-
сигеназной сигнальной системы клеток растений [Гречкин,
Тарчевский, 1999].
Абсцизовая кислота (еще один стрессовый фитогормон)
начинает интенсивно синтезироваться в клетках растений
после атаки патогенов вследствие элиситориндуцированной
экспрессии гена оксидазы зеаксантина [Audran et al., 1998;
Grill, Himmelbach,
1998]. Позднее было обнаружено стресс-
индуцированное образование одной из изоформ 9-^ыс-эпок-
сикаротиноид-диоксигеназы [Chernys, Zeevaart, 2000] - фер-
мента, регулирующего образование абсцизовой кислоты
из каротиноидов 9-цмс-виолаксантина или 9-цмс-виолаксан-
тина.
Патогениндуцированный синтез стрессового фитогор-
мона салициловой кислоты объясняется экспрессией гена
фермента бензоат-2-гидроксилазы (представителя семейства
цитохромов Р-450), катализирующего превращение бен-
зойной кислоты в салициловую [Leon et al., 1993; 1995]. При
интенсивном синтезе салицилата часть его может перево-
диться в глюкозилированную форму с помощью фермента
УДФГ: салицилат глюкозилтрансферазы. Интересно, что
быстрая экспрессия гена этого фермента вызывается экзо-
генным салицилатом и патогенами.
Как уже упоминалось выше, к числу стрессовых фито-
гормонов относят сравнительно небольшой полипептид
(названный системином), состоящий из 18 аминокислот-
ных остатков [Constabel et al., 1998]. Он был признан рядом
авторов первым идентифицированным фитогормоном по-
липептидной природы [McGurl et al., 1992; Slosarek et al.,
1995; Bergey et al., 1996; Bowles, 1998; Ryan, Pearce, 1998;
Dombrowski et al.,
1999]. Патогены, элиситоры и механи-
ческое повреждение растений вызывают интенсивную
экспрессию системина.
Рецептор системина локализован в плазмалемме [Sheer,
Ryan,
1999]. По-видимому, при посредничестве сигнальных
систем происходил системининдуцированный синтез инги-
биторов протеиназ [Slosarek et al., 1995; Dombrowski et al.,
1999; McGurl et al., 1992; 1994a],
полифенолоксидазы
[Constabel et al.,
1995] и аминопептидазы [Chao et al., 1999].
Патогениндуцированные белки, ограничивающие пита-
ние патогенов.
Одним из первых проявлений атаки патоге-
нов может считаться экскреция ими ферментов, с одной
стороны, нарушающих целостность защитных образований
растения, а с другой - обеспечивающих углеводное и азот-
ное питание патогенов. Имеются в виду кутиназы, эндопо-
лигликаназы и протеазы. Их действие на кутин кутикулы,
полисахариды и белки клеточных стенок растений приво-
дит к освобождению вторичных элиситоров - оксигениро-
ванных мономерных высокомолекулярных кислот и спир-
тов, а также олигосахаридов и олигопептидов, которые мо-
гут включать сигнальные системы клеток растений. Одной
из защитных реакций растений является синтез белковых
ингибиторов эндополигликаназ и ингибиторов протеаз
(рис. 51). Это относительно небольшие белки, подавляю-
щие активность соответствующих экскреторных фермен-
тов патогенов - грибов и бактерий.
Обнаружено, что у фасоли ген белкового ингибитора
полигалактуроназы активировался под влиянием элисито-
ров - олигогалактуронидов или глюканов [Bergmann et al.,
1994], или при механическом повреждении растений [Devoto
et al., 1998].
Возможно, что патогениндуцируемые субтилизинподоб-
ные эндопротеазы растений (в том числе кальцийактивиру-
емая) [Tornero et al., 1996; 1997; Jorda et al., 1999] также мо-
гут ограничивать питание патогенов, гидролизуя экскрети-
руемые ими ферменты (например, кутиназы, эндогликана-
зы и протеиназы) во внеклеточном пространстве.
Патоген(элиситор)индуцированные ферменты синтеза
фенилпропаноидных фитоалексинов.
Ряд патогениндуциро-
нанных белков-ферментов катализирует образование низ-
комолекулярных растительных антибиотиков - фенилпро-
маноидных или терпеноидных фитоалексинов [Метлицкий,
()зерецковская, 1985; Ebel,1986; Дмитриев, 1999] (рис. 52).
Фенилпропаноидные фитоалексины насчитывают большое
количество соединений, объединенных общностью первых
тгапов синтеза и отличающихся последними этапами. Сво-
им названием они обязаны первой реакции - образованию
фенилпропанового производного аминокислоты фенилала-
пина - коричной кислоты с помощью фермента фенил ал а-
мин-аммиак-лиазы. Насчитывается более 20 ферментов,
принимающих участие в синтезе фенилпропаноидных фи-
тоалексинов [Neumann et al., 1989]. Наиболее простыми
7-11-
углеродными продуктами превращения фенил ал анина
являются бензойная, салициловая, кумаровая, гидроксику-
маровая, кофейная, оксиметилкофейная (феруловая) кис-
лоты. Из феруловой кислоты путем гидроксилирования и
метилирования образуются 5-гидрокси-ферулат и синапо-
вая кислота. Большинство из этих соединений обладает
свойствами антибиотиков, а салициловая кислота, как уже
неоднократно упоминалось ранее, играет роль одного из
главных системных сигналов.
Важны в защите растений от грибов и бактерий более
сложные продукты фенилпропаноидного метаболизма, со-
держащие два, три, четыре и пять гетероциклов. Большин-
ство из них происходят из 15-углеродного флавоноидного
каркаса, который синтезируется с помощью халконсинтазы
из производного кумаровой кислоты - кумароил-КоА и
трех молекул малонил-КоА. Отмечена активация у люцер-
ны грибными элиситорами фермента ацетил-КоА-карбок-
силазы, катализирующей реакцию синтеза малонил-КоА из
ацетил-КоА, СО
2
и АТФ [Shorrosh et al., 1994]. Найдено, что
халконсинтаза имеет много изоформ даже в одном и том же
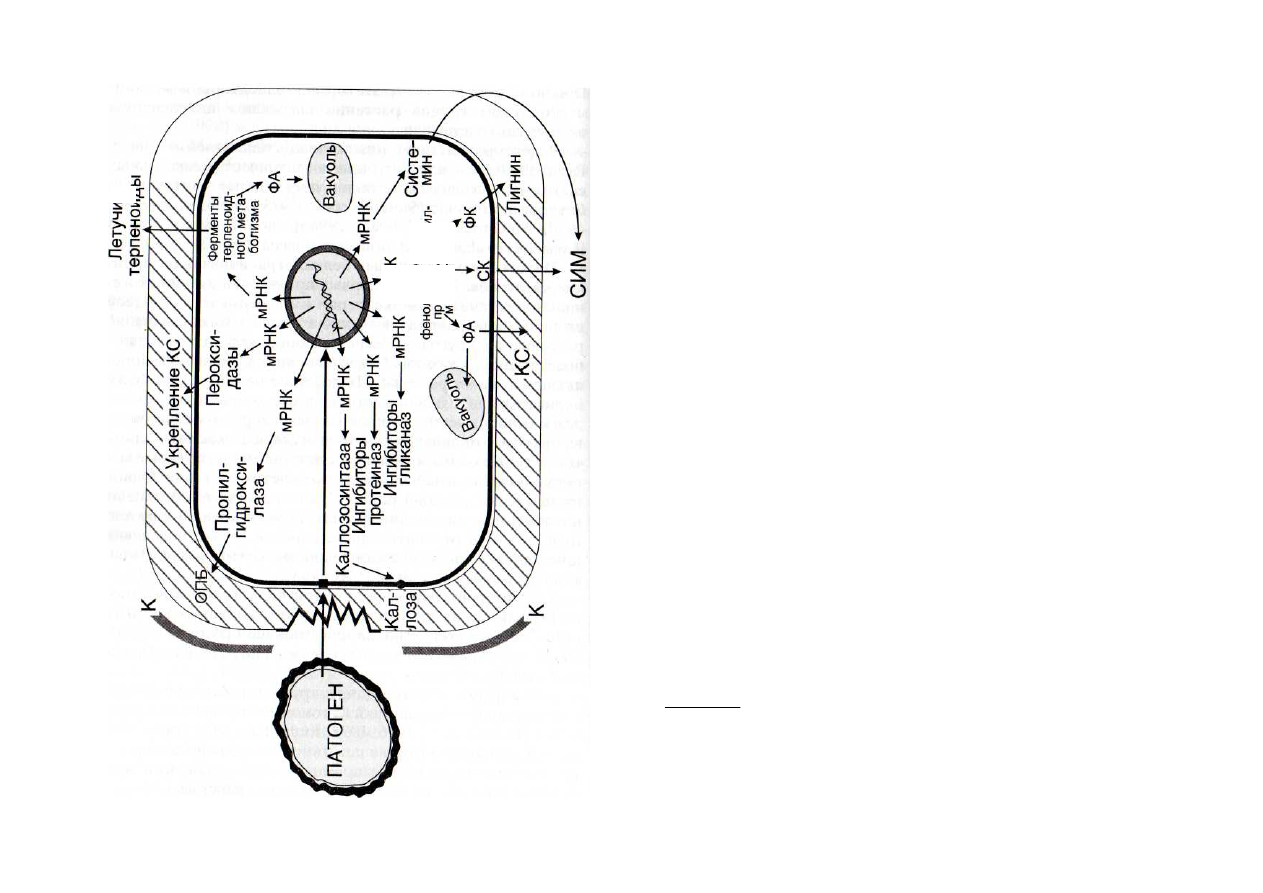
Рис. 51. Схема образования элиситориндуцируемых белков, вы-
зывающих укрепление клеточных стенок растений, синтез фито-
алексинов, системных элиситоров, ингибиторов протеиназ и гли-
каназ
К - кутикула; КС - клеточные стенки; ОПБ - оксипролиновые бел-
ки; СИМ - системный иммунитет; СК - салициловая кислота; ФА - фи-
тоалексины; ФК - фенольные кислоты
CD
ли
ъ
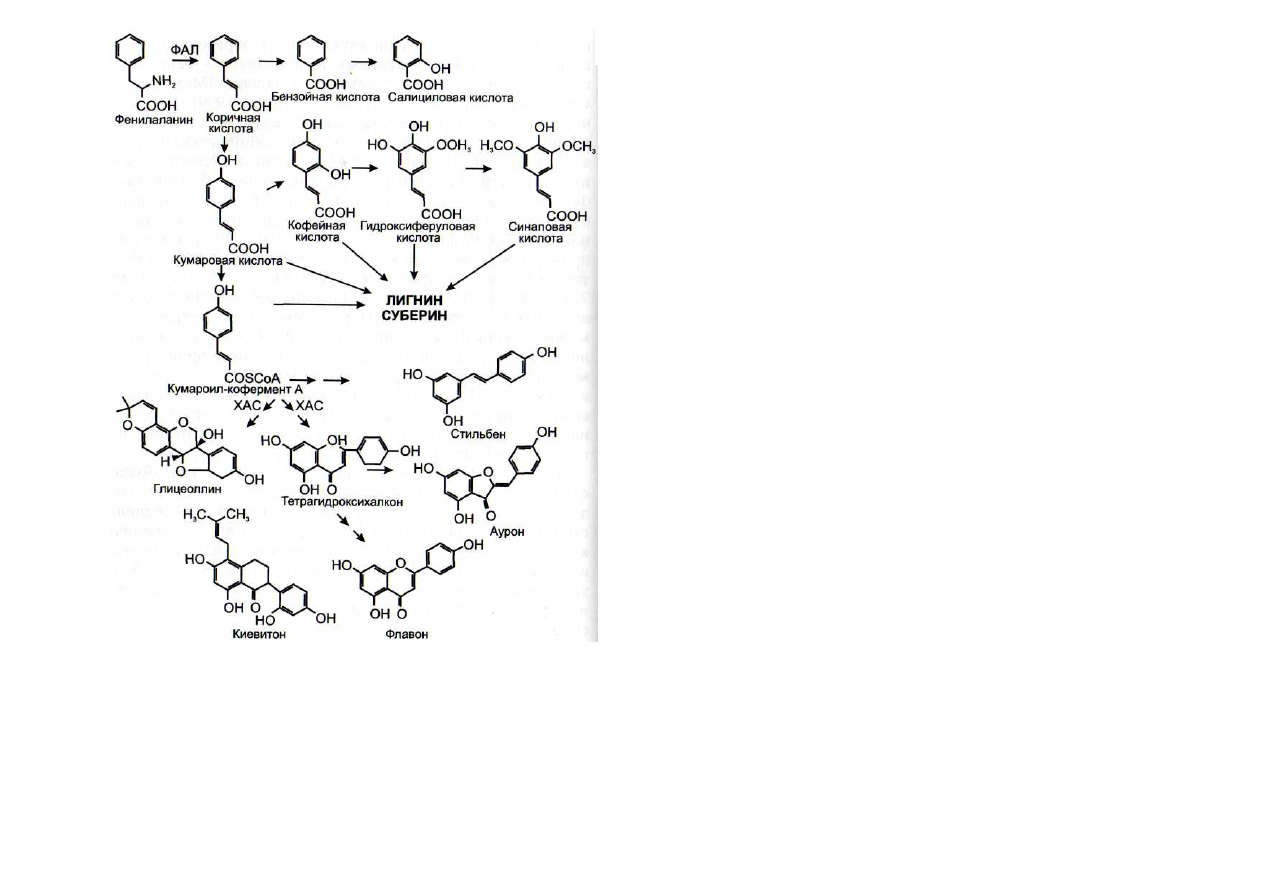
Рис. 52. Схема синтеза некоторых фенилпропаноидных фито-
алексинов (по: [Dixon, Paiva, 1995])
ФАЛ - фенилаланин-аммиак-лиаза; ХАС - халконсинтаза
растении (семь у гороха), причем они различным образом
реагируют на сигналы из окружающей среды. Так, одна из
изоформ активировалась элиситорами, но не ультрафиоле-
том, а для другой была отмечена противоположная зависи-
мость [Y. Ito et al., 1997].
Продуктом халконсинтазной реакции является тетра-
гидроксихалкон, который впоследствии превращается в
другие флавоноидные классы соединений, такие как фла-
воны, флавононы, флаванолы, антоцианины и 3-дезокси-
антоцианидины [Dixon, Paiva, 1995]. При участии халкон-
редуктазы, стильбенсинтазы, изофлавонсинтазы происхо-
дит образование различных простых изофлавоноидов, ку-
местанов, птерокарпанов и изофлаванов, которые облада-
ют ярко выраженными фунгицидной [Morrissey, Osbourn,
1999] и бактерицидной активностью, подавляя, например,
развитие стрептококков, актиномицетов, лактобацилл
[Tsuchiya et al.,
1994]. Простые фитоалексины усложняют-
ся у различных растений за счет модификационных реак-
ций гидроксилирования, гликозилирования, ацилирования,
пренилирования, сульфатации и метилирования, отража-
ющих видоспецифические особенности метаболизма, но
сохраняющих или даже усиливающих фунгицидные и бак-
терицидные свойства этих соединений. В ряде случаев по-
казано, что они эффективно защищают от нематод
[Baldridge et al., 1998].
Из различных растений выделено большое количество
индивидуальных фитоалексинов: из Brassica - брассинин,
циклобрассинин, брассилексин, из Potato - ришитин, люби-
мин, из Wasabia - вазалексины, из Arabidopsis - камалек-
син, из Orizae - момилактон и сакуранетин, из Medicago -
медикарпин, из Pisum - писатин, из Glycine - глицеоллины,
из Orchidaceae - орхинол и хирцинол, из Phaseolus - киеви-
тон, из Petroselenium - апиин, из Dianthus - диантрамидные
соединения, из Ruta - 1,3-дигидрокси-1Ч-метилакридон,
образующийся из N-метилантраноила и малонил-КоА.
Фенилпропаноидные фитоалексины - это нелетучие ве-
щества.
Образование и накопление различных фитоалексинов
происходит благодаря индукции патогенами и элиситорами
экспрессии генов, кодирующих ферменты фенилпропано-
идного метаболизма (см. рис. 52) - фенилаланин-аммиак-
лиазу [Ebel et al., 1984; Bonhoff et al., 1986; Ni et al., 1996;
Weiergang et al., 1996; Baldridge et al.,
1998], халконсинтазу
[Ebel et al., 1984; Grab et al., 1985; Bonhoff et al., 1986;
Weiergang et al., 1996; Colliver et al., 1997; Baldridge et al.,
1998; Ferrer et al.,
1999], халконизомеразу и халконредуктазу
[Ni et al.,
1996], циннамат-4-гидроксилазу [Ni et al., 1996;
Ferrer et al.,
1999], стильбенсинтазу [Bolwell, Dixon, 1986;
Tropf et al., 1994; Colliver at al.,
1997], изофлавонсинтазу
[Ebel et al.,
1984], изофлавонредуктазу [Weiergang et al.,
1996], изофлавоноидредуктазу [Tsuchiya et al.,1994], кофеат-
О-метилтрансферазу [Bonhoff et al., 1986; Tsuchiya et al.,
1994], пренилтрансферазы [Hain et al., 1993], халкон-О-ме-
тилтрансферазу [Weiergang et al., 1996], метилтрансферазу
[Hamerski et al.,
1990], акридонсинтазу [Rakwal et al., 1996],
ферменты синтеза триптофана [Junghanns et al., 1998], ак-
тивность которых коррелировала с накоплением камалек-
сина у арабидопсиса, дигидробензофенантридин-оксидазу
[Zhao, Last,
1996], ферменты семейства цитохром Р-450
[Schopfer et al.,
1998], вызывающие гидроксилирование
кольчатых фенилпропаноидных структур, и др. Было пока-
зано, что грибной элиситор вызывает неодновременное по-
вышение активности различных ферментов фенилпропано-
идного метаболизма [Chappel et al., 1984]: сначала фенил-
аланин-аммиак-лиазы, а уже затем - метилтрансферазы,
катализирующей последние этапы синтеза фенилпропано-
идных фитоалексинов.
Обнаружено, что индукция фитопатогенными грибами
синтеза фенилаланин-аммиак-лиазы у растений гороха по-
давлялась супресцином — соединением, продуцируемым гри-
бом вместе с элиситорами [Wada et al., 1995].
Патогениндуцируемые ферменты синтеза терпеноид-
ных фитоалексинов.
Для терпеноидных фитоалексинов,
так же как и для фенилпропаноидных, характерны общ-
ность начальных реакций и различия в заключительных ре-
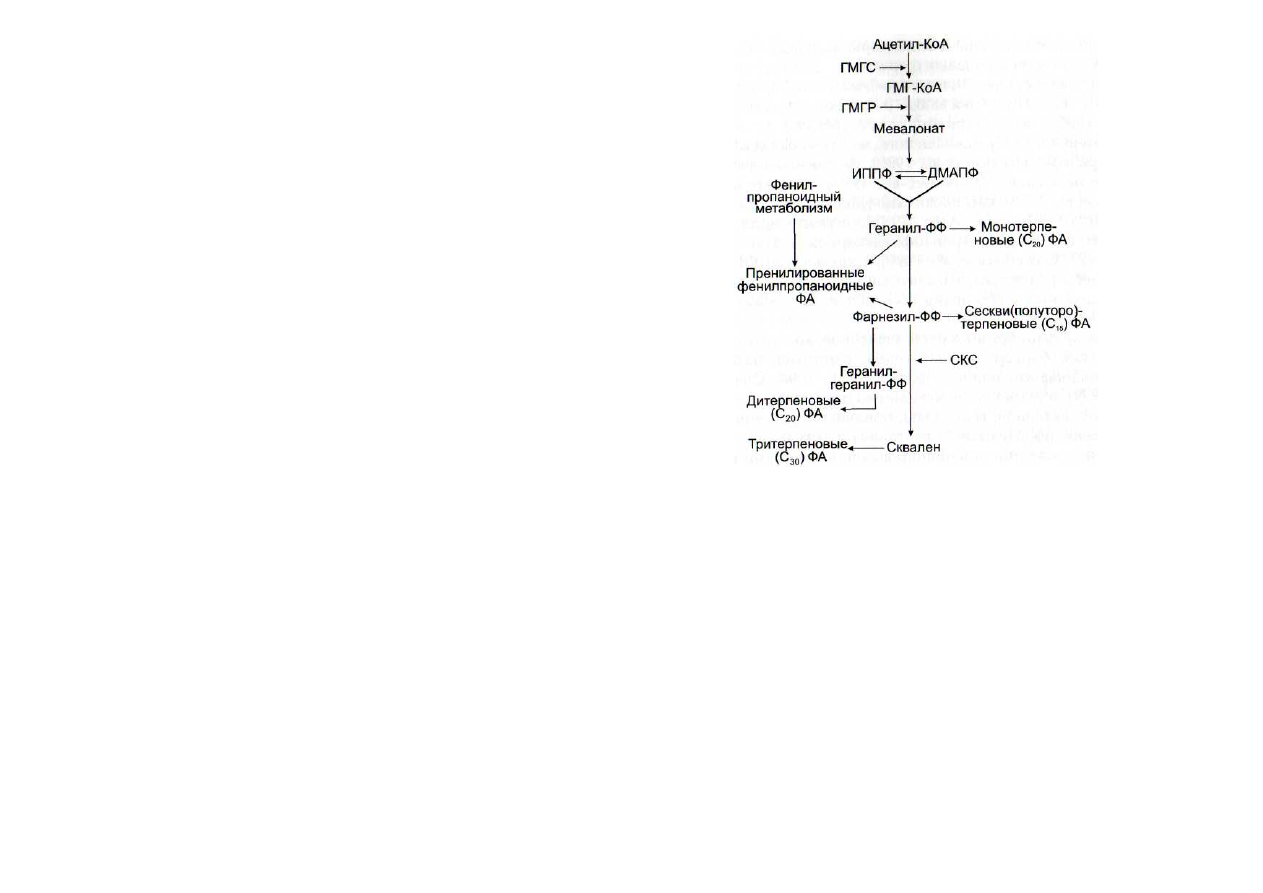
акциях метаболизма (рис. 53). Исходным субстратом их син-
теза является ацетил-КоА, промежуточными соединения-
ми - 3-гидрокси-З-метилглутарил-КоА, мевалонат, изопен-
тенилпирофосфат и диметилаллилпирофосфат, геранилпи-
рофосфат, фарнезилпирофосфат, геранилгеранилпирофос-
фат, а также сквален, который может превращаться в раз-
личные фитоалексины путем реакций циклизации и присо-
единения различных радикалов за счет реакционной спо-
собности ненасыщенных связей [Threlfall, Whitehead, 1990;
Bach,
1995]. Можно перечислить некоторых представите-
лей терпеноидных фитоалексинов: у картофеля сесквитер-
пеноидные ришитин, любимин, фитоберин, у полыни -
Рис. 53. Схема синтеза терпеноидных фитоалексинов (по:
[Threlfall, Whitehead, 1990; Bach, 1995])
ГМГС - З-гидрокси-3-метил-глутарилсинтаза; ГМГ-КоА - 3-гид-
рокси-3-метилглутарил-КоА; ГМГР - З-гидрокси-3-метилглутарил-ре-
дуктаза; ДМАПФ ~ диметилаллилпирофосфат; ИППФ - изопентенил-
пирофосфат; СКС - скваленсинтаза; ФА - фитоалексины; -ФФ - оста-
ток пирофосфата
олефиновые монотерпены лимонен, терпинолен, а- и у-
терпинены, мирцен, сесквитерпены,
а-
и (3-цедрены, (3-
фарнезен, а-акорадиен, а-бисаболен, оксигенированные
цедрол и эпицедрол, у риса - 9-(3-пимара-7,15-диен и сте-
мар-13-ен, у хлопчатника - госсипол и ласинилен, дезокси-
гемигоссипол, гемигоссипол, гемигоссиполон, гелиоциды,
у фасоли - два летучих гомотерпена 4,8-диметил-1,ЗЕ,7-ди-
метилнонатриен и 4,8,12-триметил-1,ЗЕ,7Е,11-тридекатет-
раен, у табака - капсидиол, глютинозон, ветиспирадиен, у
хиосциамус - соединения с ветиспирановым углеродным
остовом и др.
Различные патогены и элиситоры вызывают индукцию
экспрессии генов ключевых ферментов синтеза терпеноид-
ных фитоалексинов: З-гидрокси-3-метилглутарилредукта-
зы [Yang et al., 1991; Choi et al., 1992; Nelson et al., 1994; Joost
et al.,
1995], сесквитерпенциклазы [Mercke et al., 1999;
Yoshioka et al.,
1999], скваленсинтазы [Yoshioka et al., 1999],
метилтрансферазы [Liu et al., 1999], изопентенил-дифосфат-
изомеразы [Ramos-Valdivia et al., 1997], дитерпенсинтазы
[Mohan et al.,
1996], касбенсинтазы [Mohan et al., 1996], А-ка-
диенсинтазы [Moesta, West, 1985], стриктозидинсинтазы
[Chen et al.,
1995] и триптофандекарбоксилазы [Cardoso
et al., 1997; Ouwerkerk et al.,
1999], а также НАДФН: цито-
хром Р-450 редуктазы, участвующей в синтезе терпеноид-
ных индольных алкалоидов [Schopfer et al., 1998; Pasquali
et al., 1999].
К числу газообразных фитоалексинов, которые создают
химический барьер для патогенов, относятся некоторые
терпеноидные соединения [Норке et al., 1994; Ouewerkerk
et al.,
1999], а также продукты липоксигеназной сигнальной
системы: гексенали, гексенолы, ноненали и ноненолы. Эти
соединения образуются в растениях в ответ на механи-
ческое повреждение и инфицирование патогенами и могут
оказывать бактерицидное и фунгицидное действие на пато-
гены на расстоянии, еще до контакта последних с растени-
ем. Летучие продукты липоксигеназной сигнальной систе-
мы - метилжасмонат, и супероксиддисмутазной сигнальной
системы - метилсалицилат, могут выполнять роль вторич-
ных элиситоров.
Патоген(элиситор)индуцируемые белки, укрепляющие
клеточные стенки растений.
Обнаружено, что после воз-
действия элиситоров на ткани растений из них значительно
труднее получить изолированные протопласты [Bradley
et al., 1992; Brisson et al.,
1994] с помощью общепринятой ме-
тодики, предусматривающей использование целлюлаз и пек-
тиназ для разрушения клеточных стенок. Это происходит
вследствие того, что элиситоры индуцируют экспрессию
целого ряда генов, которые кодируют ферменты, катализи-
рующие образование ковалентных связей между белками
клеточных стенок, белками и полисахаридами. К числу та-
ких ферментов относятся пероксидазы [Rebmann et al., 1991;
Flocco et al.,
1998], протеиндисульфид изомераза [Esquerre-
Tugaye et al.,
1979], катализирующая образование дисуль-
фидных мостиков. В формировании более жесткой белко-
вой
структуры принимают участие не все белки клеточных
стенок, а два полипептида - 35 и 100 кДа, так как именно
они исчезают из спектра полипептидов на полиакриламид-
ном геле у вытяжек из клеточных стенок, обработанных
элиситорами растений [Bradley et al., 1992].
Усиление образования белков клеточных стенок, обога-
щенных гидроксипролином (см. рис. 51), происходит в ре-
зультате экспрессии генов пролингидроксилаз, а гликози-
лированных белков - при индукции синтеза протеин-араби-
нозил-трансфераз [Dixon et al., 1986; Corbin et al., 1987;
Denecke et al., 1995; Garcia-Muniz et al., 1998].
Укрепление клеточных стенок происходит также с по-
мощью повышения интенсивности отложения каллозы в
результате элиситориндуцированной экспрессии каллозо-
синтазы [Bonhoff et al., 1987], а также лигнина за счет
индуцированного синтеза ферментов (фенилаланин-амми-
ак-лиазы и др.) фенилпропаноидного метаболизма, обес-
печивающих образование мономерных предшественников
лигнина.
Патогениндуцируемые белки растений, вызывающие
деградацию клеточной стенки патогенов.
Одними из пер-
вых обнаруженных антипатогенных белков прямого дейст-
вия были кислые и щелочные хитиназы и р-1,3-эндоглюка-
назы [Ebel, 1986; и др.], способные гидролизовать главные
компоненты клеточной стенки грибов (рис. 54), тем самым
замедляя или прекращая рост гифов и распространение ин-
фекции [Benhamou, 1995]. В последние годы появилось до-
вольно много работ, посвященных этим ферментам, их изо-
формам, первичной структуре [Ori et al., 1990; Daugrois et al.,
1992; Buchter et al., 1997; Busam et al., 1997; Munch-Garthoff
et al., 1997; Thimmapuram et al.,
2001], промоторным участ-
кам их генов и особенностям регуляции их экспрессии
[Fukuda,
1997; Н. Wu et al., 1997], в том числе при действии
не только патогенов и элиситоров, но и различных стрессо-
вых гормонов [Simmons et al., 1992], а также механического
повреждения растений [Chang et al., 1995].
Получены данные о возможности хитиназ и (3-1,3-глю-