ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 29.06.2024
Просмотров: 512
Скачиваний: 0
СОДЕРЖАНИЕ
Более элементарные по сравнению с атоллами
Осколки частиц, или Трудное разделение
Физика возвращается к повседневным заботам
Теория наносит ответный удар: объединение
Теневая сторона стандартной модели
Проблема происхождения массы, известная как проблема полей Хиггса
Решение головоломки: как, кто, где и когда?
Предположения о происхождении жизни
Нынешняя жизнь: клеточные структуры
Решение головоломки: как, кто и почему?
Секвенирование генома человека
Решение головоломки: почему, как, кто и где, когда?
Получение или утрата атмосферного газа
Погода и климат: гипотезы (весьма добротные), прогнозы (не столь добротные)
Решение головоломки: как и где?
Измерение межзвездных расстояний
Галактики: первые теории и наблюдения
Космологический вклад Эйнштейна
Чем крупнее телескопы, тем больше расстояния до звезд
Одна большая Галактика или многочисленные обособленные галактики
Столкнувшись с неожиданным: ускорение Вселенной
В темноте рассуждать о темной энергии
Решение головоломки: где, когда, как и кто?
2. Какова доля таких звезд, имеющих планеты ?
3. Какова доля планет, обращающихся вокруг своих звезд в пределах, где возможно зарождение жизни ?
4. Какова доля благоприятно расположенных планет, где действительно зародилась жизнь?
5. Какова доля форм жизни, приведших к возникновению разума ?
7. В течение скольких лет разумная цивилизация передает в космос поддающиеся обнаружению сигналы?
13 .Предсказание землетрясений
15. Труды Эйнштейна: помимо теории относительности
Глава 2. Физика. Почему одни частицы обладают массой, а другие нет?
Глава 3. Химия. Какого рода химические реакции подтолкнули атомы к образованию первых живых существ?
Глава 4. Биология. Каково строение и предназначение протеома?
Глава 5. Геология. Возможен ли точный долговременный прогноз погоды?
Глава 6. Астрономия. Почему Вселенная расширяется со все большей скоростью?
Оперон днк — рнк — белки
Представляется, что присутствие лактозы в клетке служит пусковым механизмом, приводящим в действие транскрипцию РНК, которая производит эти три белковых фермента. На самом же деле все обстоит значительно сложнее. Сигнал к производству различных белковых ферментов задается одновременно наличием лактозы и отсутствием глюкозы. Рассмотрим этот процесс на молекулярном уровне, чтобы выяснить его механизм.
ДНК порой представляют в виде обособленной молекулы, надежно защищенной благодаря своему крепкому сложению, хранящей жизненно важную для клетки информацию. Но это далеко не так. В действительности ДНК постоянно прощупывают, крутят, тормошат, раскрывают различные белковые ферменты. Такая деятельность заставляет эту информационную магистраль изрядно выкладываться.
Все эти действия обусловлены обликом ДНК и распределением электрического заряда. Двойная спираль имеет бороздки, маленькую и большую, а все нуклеотидные основания обладают только им присущим распределением электрического заряда (см.: Список идей, 6. Сборка модели ДНК, где рассказывается, как собирать часть ДНК из набора конструктора). Некоторые белки имеют размер и очертание, приходящиеся «впору» этим бороздкам. Благодаря распределению электрического заряда у белков и ДНК они могут плотно прилегать друг к другу. Однако притяжение не столь сильно, как ковалентные связи внутри каждой молекулы. Такое вкладывание одной молекулы в другую называют связыванием.
В зависимости от формы и распределения заряда белки присоединяются в соответствующих местах вдоль ДНК. Ввиду теплового движения молекул белки постоянно связываются и отделяются.
Соответствие сложных молекулярных очертаний часто представляется аналогичным ключу и замку. Лишь немногие очертания в достаточной степени соответствуют друг другу для соединения молекул. Белки тоже могут связываться с другими белками, образуя новую единицу под названием комплекс. Обычно комплекс приобретает иные по сравнению с исходной молекулой очертание и распределение заряда. Такую перемену, играющую главную роль в сборке белка, поскольку меняются «ключи» и «замки», именуют конформационным изменением.
РНК собирается с помощью белкового фермента (полимеразы), который прикрепляется к связывающей стороне ДНК, распускает двойную спираль посередине подобно «змейке» и переписывает (транскрибирует) порядок парных оснований ДНК на молекулу РНК. Затем РНК покидает ДНК и переносит (транслирует) порядок парных нуклеотидных оснований, собирая белок на молекулярном устройстве под названием рибосома. Каждая группа из трех нуклеотидных оснований, именуемая кодоном (см.: Список идей, 7. Кодоны), определяет, какую аминокислоту добавить к белку. Полимераза РНК связывается с ДНК лишь в тех местах, где приходится впору. Это прилаживание определяется не только очертанием молекулы полимеразы, но и наличием места связывания у ДНК, которое в свою очередь зависит от изгибов ДНК.
Для получения полной картины метаболического процесса на основе лактозы недостает трех молекул. Прежде всего это белок-активатор катаболизма (БАК-белок). В обычном состоянии строение БАК-белка не позволяет ему соединяться с ДНК. БАК-белок содержит место для связывания с другой молекулой, циклическим аденозинмонофосфатом (цАМФ). Молекула цАМФ вырабатывается в среде, где отсутствует глюкоза. Если цАМФ связан с БАК-белком, БАК-белок претерпевает конформационное изменение, позволяющее ему присоединяться к ДНК. В свою очередь, связывание комплекса БАК-белок/цАМФ с ДНК Е. соli заставляет ДНК сгибаться, как показано на рис. 4.2.
На заключительном этапе требуется наличие другого белка, действующего в качестве репрессора. В данном случае он называется lас-репрессором. Эта молекула обычно входит в бороздку ДНК в том месте, где нужно помешать закрепиться полимеразе РНК, переписывающей информацию ДНК на белки, которые усваивают лактозу.
Если лактоза не соединена с lac-репрессором, репрессор точно входит в бороздку ДНК в нужном месте, препятствуя выполнению полимеразой РНК возложенной на нее задачи перезаписи (транскрипции). Если лактоза соединена с lac-репрессором, это вызовет в репрессоре конформационные изменения, так что он уже не будет подходить бороздке ДНК Е. соli и не будет мешать полимеразе ДНК выполнять транскрипцию. Рассмотрим, как эти молекулы сотрудничают, определяя наблюдаемую линию поведения Е. coli.
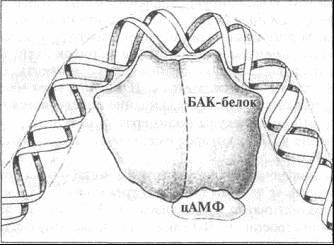
Рис. 4.2. Связь цАМФ с БАК-белком, а БАК-белка — с ДНК (из кн.: Raven P. H., Johnson G. В. Biology)
В исходных условиях имеется глюкоза и отсутствует лактоза. При наличии глюкозы цАМФ не вырабатывается, а значит, не образовывается комплекс БАК-белок/цАМФ, не сгибается ДНК и полимераза РНК не переписывает белки для усвоения лактозы. Помимо этого, репрессор находится на ДНК, препятствуя соединению полимеразы РНК с ДНК в этом месте. Получается двойная блокировка перезаписи (рис. 4.3а).
В смешанной среде с глюкозой и лактозой присутствие глюкозы препятствует образованию комплекса БАК-белок/ цАМФ, так что ДНК не изгибалась, а полимераза РНК не занималась перезаписью. Даже если присутствие лактозы вынудит репрессор покинуть бороздку ДНК, полного связывания полимеразы РНК не произойдет. Она покидает ДНК, так и не прикрепившись ни к одному из участков lac-оперона.

Рис. 4.3. Lac-penpeccop (из кн.: Raven P. H., Johnson G. В. Biology)
В отсутствие глюкозы и лактозы появляется комплекс БАК-белок/цАМФ, изгибается ДНК в ожидании РНК, но при этом наличествует и репрессор. С точки зрения Е. coli отсутствие пищи означает предстоящий голод. Но мы видим, насколько она готова к возможному повороту событий. Если появится глюкоза, она не станет расходовать энергию на выработку белковых ферментов, а сразу приступит к усваиванию глюкозы. Если же появится лактоза, изогнувшаяся ДНК уже готова к сборке нужной РНК, стоит лишь лактозе соединиться с репрессором, который тотчас покидает ДНК (рис. 4.36).
В отсутствие глюкозы и при наличии лактозы происходит следующее. Недостаток глюкозы приводит к образованию комплекса БАК-белок/цАМФ, который соединяется с ДНК, вынуждая ее изгибаться. Это дает возможность полимеразе РНК отыскать свое место прикрепления. Присутствие лактозы приводит к ее связыванию с laс-репрессором и отсоединению репрессора от ДНК, так что уже вся полимераза РНК может присоединиться к ДНК и собрать три белка для усвоения лактозы (рис. 4.3в).
Подобное положение дел сходно с ситуацией с дверью, снабженной ручкой и засовом. Ручка действует подобно активатору, а запор выступает в роли репрессора. В таблице действие системы «ручка—засов» сравнивается с механизмом «активатор—репрессор».
|
Положение ручки (активатор) |
Положение запора (репрессор) |
Откроется ли дверь (образуются ли усваивающие лактозу белки?) |
|
Не повернута (высокое содержание глюкозы) |
Заперто (высокое содержание лактозы) |
Нет |
|
Не повернута (высокое содержание глюкозы) |
Не заперто (низкое содержание лактозы) |
Нет |
|
Повернута (высокое содержание глюкозы) |
Заперто (низкое содержание лактозы) |
Нет |
|
Повернута (низкое содержание глюкозы) |
Не заперто (низкое содержание лактозы) |
Да |
Эта сложная система управления схожа со старым устройством Руба Голдберга7, где целая цепь сложных событий служит для достижения некой простой цели (рис. 4.4). И все же почему при всей затратности данного механизма Е. coli не вырабатывает постоянно все нужные ферменты, чтобы усваивать любой поступающий к ней сахар? Возможно, некогда такая бактерия и существовала.
Но появившаяся в ходе мутаций Е. coli с ее lac-опероном благодаря значительно большему по сравнению со своей старшей родственницей коэффициенту полезного действия вытеснила ее. Наглядный пример классического естественного отбора.
Белковые ферменты собираются практически одновременно с перезаписью РНК, когда РНК все еще прикреплена к длинной кольцевой молекуле ДНК. Поскольку Е. coli относится к прокариотным клеткам, у нее нет тормозящих ход метаболизма ядра или ядерной мембраны, так что усвоение лактозы начинается очень скоро. Е. coli прекрасно живет и на лактозе, и на глюкозе.
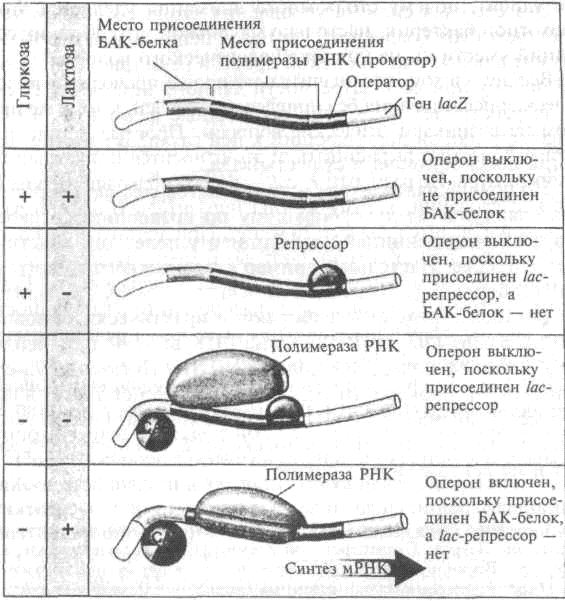
Рис. 4.4. Действие lac-оперона на молекулярном уровне (из кн.: Raven P. H., Johnson G. В. Biology)
Е. coli и другие организмы
Е. coli — одно из наиболее изученных живых существ; исследователи выявили примерно две трети функций ее генов. Механизм задействования lac-оперона составляет лишь малую часть молекулярных отправлений Е. coli. Возможно, вас удивит, почему столь много внимания уделяется этой крохотной бактерии, настолько маленькой, что 50 таких созданий уместится на кончике человеческого волоса.
Все дело в том, что значительно проще проводить биологические исследования без привлечения людей, к тому же при этом не возникают этические вопросы. Простые организмы проще и быстрее выращивать, и это относится к проведению самих опытов. По размерам Е. coli — весьма подходящий объект для исследований, к тому же она поразительно быстро размножается: делится каждые 20 минут. При достаточном количестве воды, глюкозы (или лактозы) и места за десять часов из одной Е. coli можно получить свыше 1 млрд. клеток. Если другие штаммы Е. coli опасны для здоровья, то разновидность К-12 вполне безобидна, так что нет нужды предпринимать серьезные меры предосторожности.
Вот уже свыше 70 лет Е. coli выступает рабочей лошадкой биохимии, генетики и биологии развития. Сходство ее молекулярного процессинга с происходящим в других организмах даже привело к использованию ее в качестве фабрики по производству инсулина для больных диабетом. Жак Моно, выдающийся французский ученый и Нобелевский лауреат 1965 года, однажды заметил: «Что верно для Е. coli, верно и для слона».
Изучаются и другие прокариоты, вроде Mycoplasma genitalium [возбудителя мочеполового микоплазмоза], самого крохотного самостоятельного живого организма, в составе ДНК которого 580 тыс. пар азотистых оснований и 517 генов, и Haemophilus influenzae [палочки Пфайфера, возбудителя пневмонии и гнойного менингита], в составе ДНК которого 1 830 137 пар азотистых оснований и 1743 гена. Но относительная простота прокариотной ДНК в связи с ее величиной и кольцевидностью ограничивает ее применение к более сложным организмам.