ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 12.10.2020
Просмотров: 4701
Скачиваний: 20
36
положительный заряд) и содержимым верхнего и нижнего каналов. Этот эндокохлеарный потенциал
оказывает влияние на мембранный потенциал слуховых рецепторных клеток, создавая у них
критический уровень поляризации, при котором незначительное механическое воздействие во время
контакта волосковых рецепторных клеток с текториальной мембраной приводит к возникновению в них
возбуждения.
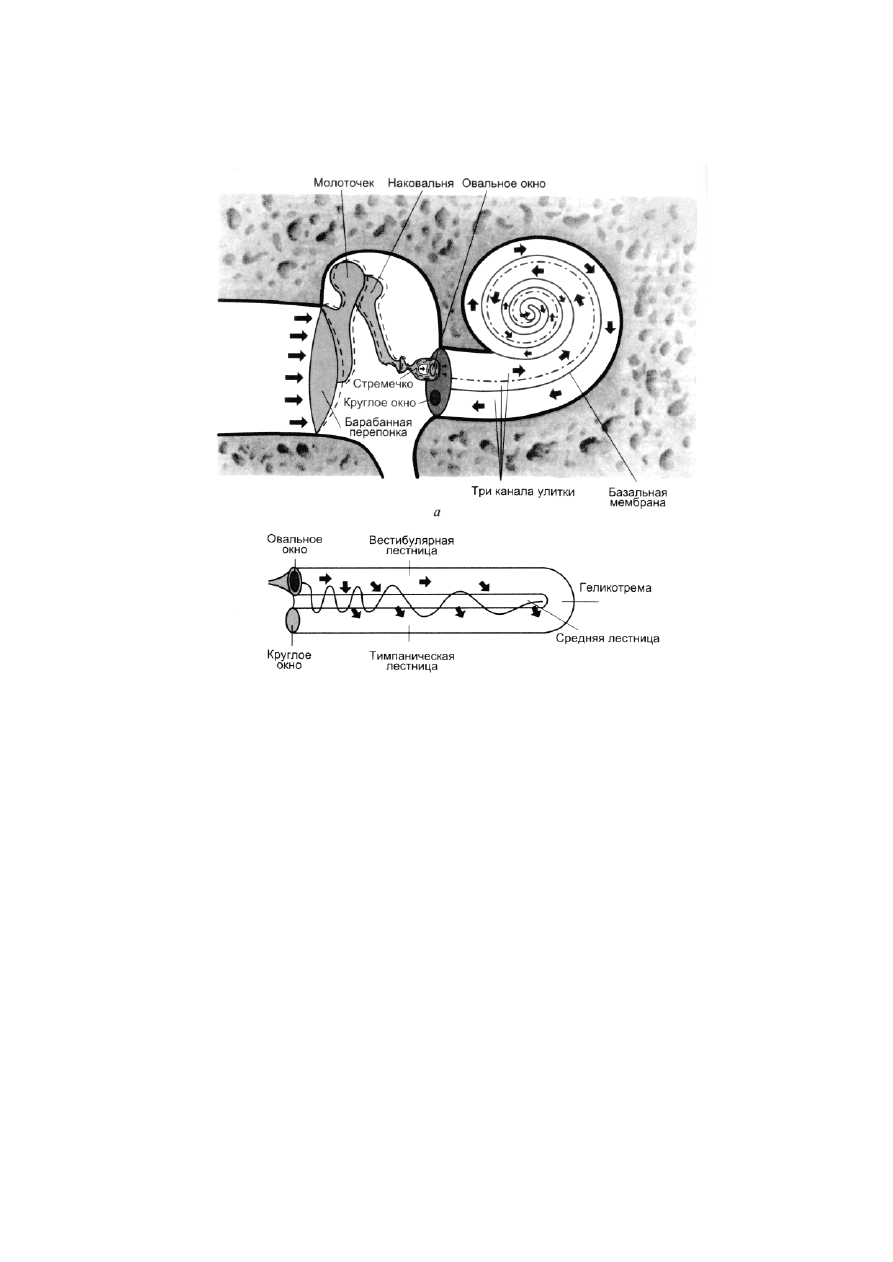
Рис. 2.8. Каналы улитки:
а
–
среднее и внутреннее ухо в разрезе (по П. Линдсею и Д. Норману, 1974);
б
–
распространение звуковых
колебаний в улитке
3. Микрофонный эффект улитки был получен в эксперименте на кошках. Электроды, введенные в
улитку, соединялись с усилителем и громкоговорителем. Если рядом с ухом кошки произносили
различные слова, то их можно услышать, находясь у громкоговорителя в другом помещении. Этот
потенциал генерируется на мембране волосковой клетки в результате деформации волосков при
соприкосновении с текториальной мембраной. Частота микрофонных потенциалов соответствует
частоте звуковых колебаний, а амплитуда потенциалов в определенных границах пропорциональна
интенсивности звуков речи. Звуковые колебания, действующие на внутреннее ухо, приводят к тому, что
возникающий микрофонный эффект накладывается на эндокохлеарный потенциал и вызывает его
модуляцию.
4. Суммационный потенциал отличается от микрофонного потенциала тем, что отражает не форму
звуковой волны, а ее огибающую. Он
представляет собой совокупность микрофонных потенциалов,
возникающих при действии сильных звуков с частотой выше 4000
–
5000 Гц. Микрофонный и
суммационный потенциалы связывают с деятельностью наружных волосковых клеток и рассматривают
как рецепторные потенциалы.
5. Потенциал действия слухового нерва регистрируется в его волокнах, частота импульсов
соответствует частоте звуковых волн, если она не превышает 1000 Гц. При действии более высоких
тонов частота импульсов в нервных волокнах не возрастает, так как
1000 имп/с
–
это почти
максимально возможная частота генерации импульсов в волокнах слухового нерва. Потенциал действия
в нервных окончаниях регистрируется через 0,5
–
1,0 мс после возникновения микрофонного эффекта,

37
что свидетельствует о синаптической передаче возбуждения с волосковой клетки на волокно слухового
нерва.
Восприятие звуков различной высоты
(частоты), согласно резонансной теории Гельмгольца,
обусловлено тем, что каждое волокно основной мембраны настроено на звук определенной частоты.
Так, звуки низкой частоты воспринимаются длинными волнами основной мембраны, расположенными
ближе к верхушке улитки, звуки высокой частоты воспринимаются короткими волокнами основной
мембраны, расположенными ближе к основанию улитки. При действии сложного звука возникают
колебания различных волокон мембраны.
В современной интерпретации резонансный механизм лежит в основе
теории места,
в соответствии
с которой в состояние колебания вступает вся мембрана. Однако максимальное отклонение основной
мембраны улитки происходит только в определенном месте. При увеличении частоты звуковых
колебаний максимальное отклонение основной мембраны смещается к основанию улитки, где
располагаются более короткие волокна основной мембраны,
–
у коротких волокон возможна более
высокая частота колебаний. Возбуждение волосковых клеток именно этого участка мембраны при
посредстве медиатора передается на волокна слухового нерва в виде определенного числа импульсов,
частота следования которых ниже частоты звуковых волн (лабильность нервных волокон не превышает
800 –
1000 Гц). Частота воспринимаемых звуковых волн достигает 20 000 Гц. Таким способом
осуществляется пространственный тип кодирования высоты и частоты звуковых сигналов.
При действии тонов примерно до 800 Гц кроме
пространственного
кодирования происходит еще и
временное (частотное)
кодирование, при котором информация передается также по определенным
волокнам слухового нерва, но в виде импульсов (залпов), частота следования которых повторяет
частоту звуковых колебаний. Отдельные нейроны на разных уровнях слуховой сенсорной системы
настроены на определенную частоту звука, т.е. каждый нейрон имеет свой специфический частотный
порог, свою определенную частоту звука, на которую реакция нейрона максимальна. Таким образом,
каждый нейрон из всей совокупности звуков воспринимает лишь определенные достаточно узкие
участки частотного диапазона, не совпадающие между собой, а совокупности нейронов воспринимают
весь частотный диапазон слышимых звуков, что и обеспечивает полноценное слуховое восприятие.
Правомерность этого положения подтверждается результатами протезирования слуха человека, когда
электроды вживлялись в слуховой нерв, а его волокна раздражались электрическими импульсами
разных частот, которые соответствовали звукосочетаниям определенных слов
и фраз, обеспечивая
смысловое восприятие речи.
Анализ интенсивности звука
также осуществляется в слуховой сенсорной системе. При этом сила
звука кодируется как частотой импульсов, так и числом возбужденных рецепторов и соответствующих
нейронов. В частности, наружные и внутренние волосковые рецепторные клетки имеют разные пороги
возбуждения. Внутренние клетки возбуждаются при большей силе звука, чем наружные. Кроме того, у
внутренних клеток пороги возбуждения также различны. В связи с этим в зависимости от
интенсивности звука меняются соотношение возбужденных рецепторных клеток кортиева органа и
характер импульсации, поступающей в ЦНС. Нейроны слуховой сенсорной системы имеют различные
пороги реакций. При слабом звуковом сигнале в реакцию вовлекается лишь небольшое число более
возбудимых нейронов, а при усилении звука возбуждаются нейроны с меньшей возбудимостью.
Необходимо отметить, что кроме воздушной проводимости имеется
костная проводимость звука,
т.е. проведение звука непосредственно через кости черепа. При этом звуковые колебания вызывают
вибрацию костей черепа и лабиринта, что приводит к повышению давления перилимфы в
вестибулярном канале больше, чем в барабанном, так как перепонка, закрывающая круглое окно,
эластична, а овальное окно закрыто стремечком. В результате этого происходит смещение основной
мембраны, так же как и при воздушной передаче звуковых колебаний.
Определение
локализации источника звука
возможно с помощью
бинаурального слуха,
т. е.
способности слышать одновременно двумя ушами. Благодаря бинауральному слуху человек способен
более точно локализовать источник звука, чем при монауральном слухе, и определять направление
звука. Для высоких звуков определение их источника обусловлено разницей силы звука, поступающего
к обоим ушам, вследствие различной их удаленности от источника
звука. Для низких звуков важной
является разность во времени между приходом одинаковых фаз звуковой волны к обоим ушам.
Определение местоположения звучащего объекта осуществляется либо путем восприятия звуков
непосредственно от звучащего объекта
–
первичная локализация, либо путем восприятия отраженных
от объекта звуковых волн
–
вторичная локализация, или эхолокация. При помощи эхолокации

38
ориентируются в пространстве некоторые животные (дельфины, летучие мыши).
Слуховая адаптация
–
это изменение слуховой чувствительности в процессе действия звука. Она
складывается из соответствующих изменений функционального состояния всех отделов слухового
анализатора. Ухо, адаптированное к тишине, обладает более высокой чувствительностью к звуковым
раздражениям (слуховая сенситизация). При длительном слушании слуховая чувствительность
снижается. Большую роль в слуховой адаптации играет ретикулярная формация, которая не только
изменяет активность проводникового и коркового отделов слухового анализатора, но и за счет
центробежных влияний регулирует чувствительность слуховых рецепторов, определяя уровень их
«настройки» на восприятие слуховых раздражителей.
2.3. Вестибулярный и двигательный (кинестетический) анализаторы
2.3.1. Вестибулярный
анализатор
Вестибулярный анализатор обеспечивает так называемое акселерационное чувство, т.е. ощущение,
возникающее при прямолинейном и вращательном ускорении движения тела, а также при изменениях
положения головы. Вестибулярному анализатору принадлежит ведущая роль в пространственной
ориентации человека, сохранении его позы.
Структурно
-
функциональная
характеристика.
Периферический
(рецепторный)
отдел
вестибулярного
анализатора
представлен
волосковыми
клетками
вестибулярного
органа,
расположенного, как и улитка, в лабиринте пирамиды височной кости. Вестибулярный орган (орган
равновесия, орган гравитации) состоит из трех полукружных каналов и преддверия (рис. 2.9).
Полукружные каналы расположены в трех взаимно перпендикулярных плоскостях: верхний
–
во
фронтальной, задний
–
в сагиттальной и наружный
–
в горизонтальной. Преддверие состоит из двух
мешочков: круглого (саккулюс), расположенного ближе к улитке, и овального (утрикулюс),
расположенного ближе к полукружным каналам. Полукружные каналы своими устьями открываются в
преддверие и сообщаются с ним пятью отверстиями(колено двух каналов, а именно верхнего и заднего,
соединены вместе). Один конец каждого канала имеет расширение, которое называется ампулой. Все
эти структуры состоят из тонких перепонок и образуют перепончатый лабиринт, внутри которого
находится эндолимфа. Вокруг перепончатого лабиринта и между ним и его костным футляром имеется
перилимфа, которая переходит в перилимфу органа слуха. В каждом мешочке преддверия имеются
небольшие возвышения, называемые пятнами, а в ампулах полукружных каналов
–
гребешками. Они
состоят из нейроэпителиальных клеток, имеющих на свободной поверхности волоски (реснички),
которые разделяются на две группы: тонкие, их много,
–
стереоцилии и один более толстый и длинный
на периферии пучка
–
киноцилии (рис. 2.10).
Рис. 2.9. Вестибулярный орган
Волосковые клетки представляют собой рецепторы вестибулярного анализатора и являются
вторичными. Рецепторные клетки преддверия покрыты желеобразной массой, которая состоит в
основном из мукополисахаридов, благодаря содержанию значительного количества кристаллов
карбоната кальция получила название
отолитовой мембраны.
В ампулах полукружных каналов

39
желеобразная масса не содержит солей кальция и называется
листовидной мембраной
(купулой).
Волоски рецепторных клеток пронизывают эти мембраны. Возбуждение волосковых клеток происходит
вследствие скольжения мембраны по волоскам, изгибания волосков (стереоцилии) в сторону
киноцилии. При этом возникает рецепторный потенциал волосковых клеток и
выделяется медиатор
ацетилхолин, который стимулирует синаптические окончания волокон вестибулярного нерва. Этот
эффект проявляется в усилении постоянной спонтанной активности вестибулярного нерва. Если же
смещение стереоцилии направлено в противоположную от киноцилии сторону, то спонтанная
активность вестибулярного нерва снижается.
Рис. 2.10. Структурно
-
функциональные элементы волосковой (рецепторной) клетки вестибулярного аппарата
Для
волосковых клеток преддверия
адекватными раздражителями являются ускорение или
замедление прямолинейного движения тела, а также наклоны головы. Под действием ускорения
отолитовая мембрана скользит по волосковым клеткам, а при изменении положения головы меняет свое
положение по отношению к ним. Это вызывает отклонение ресничек и возникновение возбуждения в
рецепторных волосковых клетках. Порог различения ускорения равен 2
–
20 см/с. Порог различения
наклона головы в сторону составляет около 1°, а вперед и назад
–
около 2°. При сопутствующих
раздражениях (вибрация, качка, тряска) происходит снижение чувствительности вестибулярного
аппарата. Так, вибрации, имеющие место в самолетах, повышают порог различения наклона головы
вперед и назад до 5°, при наклонах в стороны
–
до 10°.
Для
волосковых клеток полукружных каналов
адекватным раздражителем является ускорение или
замедление вращательного движения в какой
-
либо плоскости. Поскольку полукружные каналы
заполнены эндолимфой, имеющей такую же плотность, как и купула ампул, линейные ускорения не
оказывают влияния на положение ресничек и купулы. При поворотах головы или вращении тела, т.е.
при появлении углового ускорения, эндолимфа в них в силу своей инерции в первый момент остается
неподвижной или потом движется, но с иной скоростью, нежели полукружные каналы. Это вызывает
сгибание
ресничек рецепторов в купуле и возбуждение их. В зависимости от характера вращательного
ускорения или замедления происходит неодинаковое раздражение рецепторов различных полукружных
каналов. По картине импульсов, приходящих в центральные структуры вестибулярного анализатора из
полукружных каналов с каждой стороны, мозг получает информацию о характере вращательного
движения. Рецепторы полукружных каналов дают возможность различать угловое ускорение, равное в
среднем 2
–
3°/с (порог различения вращения).
40
Проводниковый отдел.
К рецепторам подходят периферические волокна биполярных нейронов
вестибулярного ганглия, расположенного во внутреннем слуховом проходе (первый нейрон). Аксоны
этих нейронов в составе вестибулярного нерва направляются к вестибулярным ядрам продолговатого
мозга (второй нейрон). Вестибулярные ядра продолговатого мозга (верхнее
–
ядро Бехтерева,
медиальное
–
ядро Швальбе, латеральное
–
ядро Дейтерса и нижнее
-
ядро Роллера) получают
дополнительную информацию по афферентным нейронам от проприорецепторов мышц или от
суставных сочленений шейного отдела позвоночника. Эти ядра вестибулярного анализатора тесно
связаны с различными отделами центральной нервной системы. Благодаря этому обеспечиваются
контроль и управление эффекторными реакциями соматического, вегетативного и сенсорного
характера. Третий нейрон расположен в ядрах зрительного бугра, откуда возбуждение направляется в
кору полушарий.
Центральный отдел
вестибулярного анализатора локализуется в височной области коры большого
мозга, несколько кпереди от слуховой проекционной зоны (21
–
22 поля по Бродману, четвертый
нейрон).
Функциональные связи вестибулярного анализатора.
При возбуждении вестибулярного
анализатора
возникают
соматические
реакции,
которые
осуществляются
благодаря
вестибулоспинальным связям при участии вестибулоретикулярных и вестибулоруброспинальных
трактов. При этом происходят перераспределение тонуса скелетной мускулатуры и рефлекторные
реакции, необходимые для сохранения равновесия тела в пространстве. Рефлексы, обеспечивающие
данную функцию, подразделяются на две группы
–
статические и статокинетические.
Один из статокинетических рефлексов
–
вестибулярный нистагм (головы или глаз)
–
имеет большое
клиническое значение. Нистагм возникает в условиях быстрого перемещения тела или его вращения.
Так, глазной нистагм проявляется сначала в ритмическом медленном движении глаз в сторону,
противоположную вращению, а затем
–
быстром движении глаз (скачком) в обратном направлении.
Реакции такого типа обеспечивают возможность обзора пространства в условиях перемещения тела.
Важным моментом является связь вестибулярного аппарата с мозжечком, благодаря чему
осуществляется тонкая регуляция моторных вестибулярных рефлексов. При нарушениях функции
мозжечка эти рефлексы утрачивают тормозной компонент, что проявляется в возникновении таких
симптомов, как, например, спонтанно возникающий нистагм, утрата равновесия, избыточная амплитуда
движений. Эти симптомы являются частью синдрома мозжечковой атаксии. Благодаря связям
вестибулярных ядер с вегетативной нервной системой проявляются вестибуловегетативные реакции
сердечно
-
сосудистой системы, желудочно
-
кишечного тракта и других органов. Они могут проявляться
в изменениях сердечного ритма, тонуса сосудов, артериального давления, усилении моторики желудка
и кишечника, повышении саливации, тошноте, рвоте и т.д. В условиях невесомости (в космосе)
возникает такой тип афферентной импульсации с вестибулярного аппарата, который никогда не
встречается на Земле. Однако привыкание к условиям невесомости во время космических полетов
происходит быстро. При этом следует учитывать, что космонавты проходят напряженный курс
тренировки, чем и объясняется их малая подверженность влиянию условий невесомости.
2.3.2. Двигательный (кинестетический) анализатор
Двигательный, или кинестетический (проприоцептивный), анализатор обеспечивает формирование
так называемого мышечного чувства при изменении напряжения мышц, их оболочек, суставных сумок,
связок, сухожилий. В мышечном чувстве можно выделить три составляющих: чувство положения,
когда
человек может определить положение своих конечностей и их частей относительно друг друга; чувство
движения, когда, изменяя угол сгибания в суставе, человек осознает скорость и направление движения;
чувство силы, когда человек может оценить мышечную силу, нужную для движения или удерживания
суставов в определенном положении при подъеме или перемещении груза. Наряду с кожным,
зрительным, вестибулярным двигательный анализатор оценивает положение тела в пространстве, позу,
участвует в координации мышечной деятельности.
Периферический отдел
представлен проприорецепторами, расположенными в мышцах, связках,
сухожилиях, суставных сумках, фасциях. К ним относятся мышечные веретена, тельца Гольджи, тельца
Пачини, свободные нервные окончания.
Мышечное веретено
представляет собой скопление тонких коротких поперечно
-
полосатых
мышечных волокон, которые окружены соединительнотканной капсулой. Эти волокна получили