ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 22.11.2023
Просмотров: 553
Скачиваний: 3
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
, что делает эти отложения очень важными в палеоботанике. Считается, что представители этого отдела являются первенцами сухопутных растений.
Риниофиты необычны тем, что прекрасно сохранились в очень древних окаменелостях и являются самыми ранними известными сосудистыми растениями. Данные окаменелостей содержат убедительные детали, подтверждающие наличие ребристого стебля, покрытого чешуевидными придатками, а также крупных двойных спорангиев. Окаменелости содержат следы спорофитов и гаметофитов других растений, остатки животных, например, членистоногих, которые обитали в одной экосистеме с риниофитами.
Известно, что риниофиты не имели полноценных корней, вместо них — корневищеподобные образования, покрытые ризоидами.
Жизненный цикл высших растений состоит из двух фаз (или поколений) – бесполой, или спорофазы (спорофита), и половой, или гаметофазы (гаметофита). у растений возможно два способа размножения – половой (с помощью гамет) и бесполый (с помощью спор), соответственно этим способам называют сами поколения.
Как половая, так и бесполая формы размножения имеют определенные биологические преимущества. При половом размножении достигается комбинация наследственного материала родительских форм. Образующаяся при половом воспроизведении особь генетически не тождественна ни одному из своих родителей. Половой процесс обеспечивает рекомбинативную генетическую изменчивость организмов из поколения в поколение, поэтому половое размножение дает виду преимущества, реализующиеся при естественном отборе. При бесполом размножении наследственные особенности передаются без изменений и могут легко закрепляться в ряду поколений, при этом как бы «тиражируется» наследственный материал родительской особи и возможен быстрый рост численности. Однако чаще всего у растений осуществляется именно чередование полового и бесполого поколений. для высших растений характерно гетероморфное чередование поколений. Оба поколения развиваются либо независимо как самостоятельные особи, либо одно поколение не образует самостоятельного растения
, а «поселяется» на другом. Например, у мхов спорофит развивается на зеленом гаметофите, а у семенных растений очень сильно редуцированный бесхлорофилльный гаметофит никогда не покидает спорофита, находясь по существу в сформированных спорофитом органах. У всех высших растений, кроме мохообразных, в жизненном цикле преобладает спорофит, гаметофит развит слабее и относительно недолговечен. В жизненном цикле мохообразных преобладает гаметофит
В эволюции остальных высших растений происходит постепенная редукция (уменьшение и упрощение) гаметофита и преобладание в жизненном цикле спорофита. Среди причин, которые обобусловили преобладание спорофита, можно назвать его диплоидность. Она по сравнению с гаплоидностью обеспечивает более высокий уровень синтетических процессов, а, с другой стороны, рецессивные мутации, снижающие жизнеспособность организма, при диплоидном состоянии не проявляются в фенотипе, то есть генотипически диплоидный организм можно считать более «стабильным».
Отдел Печёночники и мхи – Bryophyta (25 тыс. видов)
ОтделBryophyta – наиболее примитивный из всех групп наземных растений. В этот отдел входят два главных класса – Hepaticae (печеночники) и Musci (мхи). Произошли от псилофитов в силурийском периоде палеозойской эры.
Систематика и основные признаки Bryophyta
1.Чередование поколений, при котором преобладает гаметофитное поколение
2.Нет проводящей ткани, т.е. нет ни ксилемы, ни флоэмы
3.Тело представлено талломом (слоевищем) или слегка дифференцировано на простые «листья» и «стебли».
4.Нет настоящих корней, стеблей и листьев: гаметофит прикрепляется к субстрату нитевидными ризоидами. Вода и минеральные соли поглощаются всей поверхностью тела, в том числе и ризоидами. А это значит, что ризоиды – не настоящие корни, они лишь закрепляют растение в грунте. (В настоящих корнях, как и в настоящих стеблях или листьях имеются проводящие ткани). Поэтому стебли и листья мохообразных не гомологичны стеблям и листьям сосудистых растений, у которых эти органы входят в состав диплоидного спорофита, а не гаплоидного гаметофита.
Поможем написать работу на аналогичную тему
5. Спорофит прикреплён к гаметофиту, полностью зависит от него и питается за его счёт
6. Споры на спорофите образуются в споровой коробочке, расположенной на конце тонкой ножки, возвышающейся над гаметофитом
7. Встречаются главным образом в сырых затенённых местах. Могут выдерживать периоды засухи, используя для этой цели какие-то не совсем понятные механизмы.
Чередование поколений
Листостебельное фотосинтезирующее тело мха представляет собой гаметофит. На нём развиваются антеридии (мужские половые органы) и архегонии (женские половые органы). В антеридиях образуются двужгутиковые сперматозоиды, в архегониях – крупные яйцеклетки. При помощи воды (в сырую погоду) сперматозоиды выходят из антеридиев. Один из них проникает в архегоний и оплодотворяет яйцеклетку. Из образовавшейся зиготы вырастает бесполое поколение – спорофит. Он представлен коробочкой на ножке, которая вырастает на гаметофите. Спорофит не способен к фотосинтезу и питается за счёт гаметофита. В коробочке образуются гаплоидные споры. При прорастании споры формируется протонема – тонкая разветвлённая нить, на которой образуются почки, дающие начало новым гаметофитам.
Цикл развития мха
отдел моховидные
Болота сосредоточены в основном в лесной зоне северного полушария, а также во влажных экваториальных районах Африки и Южной Америки. Общая площадь болот в мире – 350 млн га. На территории нашей страны сосредоточено около 3/4 мировых болот. Основной фон растительности верховых болот составляют сфагновые мхи.
В мире насчитывается 25 тысяч видов мохообразных, из них 15–20 тысяч видов листостебельных (зеленых и сфагновых) мхов. По количеству видов среди высших растений мхи занимают второе место после цветковых (покрытосеменных). Моховидные произошли 350 млн лет назад от первых наземных растений – псилофитов, потомков прибрежных водорослей. Это многолетние растения, обычно низкорослые, их размеры от 1 мм до нескольких см; они имеют стебель, листья, корней нет. Обитают во влажных местах.
Мхи отличаются простой внутренней организацией. В их теле можно обнаружить ассимиляционную (основную) ткань, содержащую хлоропласты, а также слабо выраженные проводящие, механические, запасающие и покровные ткани. Мхи прикрепляются к почве тонкими нитевидными одноклеточными или многоклеточными выростами, расположенными на нижней части стебля, ризоидами, через которые всасываются питательные вещества из почвы.
В отличие от настоящих корней многоклеточные ризоиды состоят из одинаковых клеток и лишены проводящих тканей.
Класс печеночники - hepaticopsida. Общее число видов около 10 000 (235 родов). Их подразделяют на 7 порядков. Важнейшие из них следующие: Калобриальные -- Calabryales, Юнгерманиальные -- Jungermanniales, Маршанциальные -- Marchantiales, Сферокарпальные -- Sphaerocarpales, Риччиаль-ные -- Ricctales.
По данным морфологического и ботанико-географического анализов, виды печеночников, равно как и антоцеротовых, очень древние. Здесь явно господствуют монотипные или олиготипные роды, а иногда и семейства Из числа немногих политипных таксонов печеночников необходимо отметить лишь некоторые семейства и роды юнгерманиальных. Полиморфизм юнгерманиальных свидетельствует об их относительной молодости и процветании. Значит, наряду с архаичными, реликтовыми, т. е. угасающими таксонами, представлены и развивающиеся.
В целом же юнгерманиальные подразделяют на 25 семейств, включающих 180 родов. Общее число видов превышает 8000, что составляет около 80 % всех печеночников.
По данным палеонтологии, наиболее примитивные (талломные) формы печеночников нужно отнести к девону. Более совершенные формы, близкие к маршанциальным, найдены в пермских отложениях. Типичные маршанциальные обнаружены в начале юрского периода. Юнгерманиальные--наиболее молодая группа; ископаемые остатки известны с конца мезозойской эры.
В географическом распространении печеночники практически не имеют границ. Класс в целом характеризуется как космополитический. При этом нужно отметить, что некоторые роды и виды имеют дизъюнктивное распространение, т. е. встречаются как бы разорвано -- на разных континентах: в Южной Америке, Австралии, Южной Африке, Юго-Восточной Индии.
Такой характер распространении неоспоримо свидетельствует о большой древности этих групп растений. На древность печеночников указывает и примитивная слоевищная структура тела и число хромосом (п = 4). Такой же набор хромосом имеют и некоторые зеленые водоросли. Однако слоевище у печеночников высоко специализировано.
В качестве представителя печеночников рассмотрим маршанцию. Как и некоторые другие печеночные мхи, маршанция -- исключительно слоевищное растение (рис. 3).
Вегетативное тело спинно-брюшной симметрии, пластинчатое, дихотомически ветвящееся, 10--12 см и длину. На морфологически верхней стороне таллома развиваются на ножках особые образования -- подставки, а на них -- органы полового размножения. На одних экземплярах подставки имеют форму девятилучевой звездочки, между лучами которой на нижней стороне расположены архегонии. На других экземплярах вырастают мужские подставки, имеющие форму восьмиугольных дисков или щитков. Антеридии погружены в литеридиальные полости на верхней стороне диска -- подставки.
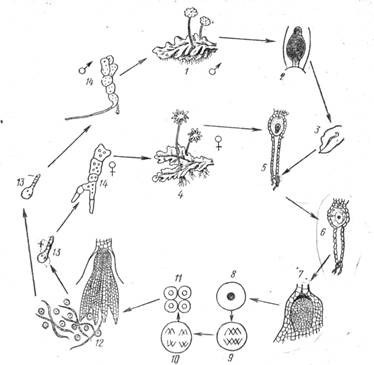
Рис. 3. Цикл развития маршанции: 1 -- мужской гаметофит, 2--антеридий, 3 -- сперматозоид, 4 -- женский гаметофит, 5 -- архегоннй, 6 -- зигота, 7 -- начальная стадия развития спорогона, 8 -- спорогенная клетка, 9-- развитие из спорогенной клетки четырех спор путем мейоза, 12 -- раскрывшийся спороген, 13--прорастание споры, 14 -- молодой таллом
Макро- и микроскопическое строение пластинчатого таллома печеночников довольно сложное. С обеих сторон находится однослойная эпидерма, а внутри--крупноклеточная бесцветная паренхима и хлорофиллоносные клетки. Верхняя эпидерма несет вентиляционные отверстия-- устьица, образованные бесцветными клетками, расположенными друг над другом в четыре ряда. Под эпидермой находятся воздушные камеры. На дне этих камер развиваются нитевидные выросты-- ассимиляторы. Они состоят из хлорофиллоносных клеток. От нижней эпидермы отходят выросты, четко различающиеся между собой. Одни из них бесцветные одноклеточные ризоиды -- простые, и язычковые. Другие, более или менее окрашенные н(красноватые или зеленоватые),-- многоклеточные чешуйки, именуемые амфигастриями. Бриологи считают, что амфигастрии представляют собой видоизмененные и редуцированные «листья» -- филлоиды. Это дает основание полагать, что слоевищные печеночники (типа маршанции) произошли от « листостебельных».
Риниофиты необычны тем, что прекрасно сохранились в очень древних окаменелостях и являются самыми ранними известными сосудистыми растениями. Данные окаменелостей содержат убедительные детали, подтверждающие наличие ребристого стебля, покрытого чешуевидными придатками, а также крупных двойных спорангиев. Окаменелости содержат следы спорофитов и гаметофитов других растений, остатки животных, например, членистоногих, которые обитали в одной экосистеме с риниофитами.
Известно, что риниофиты не имели полноценных корней, вместо них — корневищеподобные образования, покрытые ризоидами.
-
Характеристика циклов воспроизведения высших растений. Равно- и разноспоровость. Направления эволюции высших растений.
Жизненный цикл высших растений состоит из двух фаз (или поколений) – бесполой, или спорофазы (спорофита), и половой, или гаметофазы (гаметофита). у растений возможно два способа размножения – половой (с помощью гамет) и бесполый (с помощью спор), соответственно этим способам называют сами поколения.
Как половая, так и бесполая формы размножения имеют определенные биологические преимущества. При половом размножении достигается комбинация наследственного материала родительских форм. Образующаяся при половом воспроизведении особь генетически не тождественна ни одному из своих родителей. Половой процесс обеспечивает рекомбинативную генетическую изменчивость организмов из поколения в поколение, поэтому половое размножение дает виду преимущества, реализующиеся при естественном отборе. При бесполом размножении наследственные особенности передаются без изменений и могут легко закрепляться в ряду поколений, при этом как бы «тиражируется» наследственный материал родительской особи и возможен быстрый рост численности. Однако чаще всего у растений осуществляется именно чередование полового и бесполого поколений. для высших растений характерно гетероморфное чередование поколений. Оба поколения развиваются либо независимо как самостоятельные особи, либо одно поколение не образует самостоятельного растения
, а «поселяется» на другом. Например, у мхов спорофит развивается на зеленом гаметофите, а у семенных растений очень сильно редуцированный бесхлорофилльный гаметофит никогда не покидает спорофита, находясь по существу в сформированных спорофитом органах. У всех высших растений, кроме мохообразных, в жизненном цикле преобладает спорофит, гаметофит развит слабее и относительно недолговечен. В жизненном цикле мохообразных преобладает гаметофит
В эволюции остальных высших растений происходит постепенная редукция (уменьшение и упрощение) гаметофита и преобладание в жизненном цикле спорофита. Среди причин, которые обобусловили преобладание спорофита, можно назвать его диплоидность. Она по сравнению с гаплоидностью обеспечивает более высокий уровень синтетических процессов, а, с другой стороны, рецессивные мутации, снижающие жизнеспособность организма, при диплоидном состоянии не проявляются в фенотипе, то есть генотипически диплоидный организм можно считать более «стабильным».
-
Общая характеристика и принципы классификации отдела Bryophyta.
Отдел Печёночники и мхи – Bryophyta (25 тыс. видов)
ОтделBryophyta – наиболее примитивный из всех групп наземных растений. В этот отдел входят два главных класса – Hepaticae (печеночники) и Musci (мхи). Произошли от псилофитов в силурийском периоде палеозойской эры.
Систематика и основные признаки Bryophyta
1.Чередование поколений, при котором преобладает гаметофитное поколение
2.Нет проводящей ткани, т.е. нет ни ксилемы, ни флоэмы
3.Тело представлено талломом (слоевищем) или слегка дифференцировано на простые «листья» и «стебли».
4.Нет настоящих корней, стеблей и листьев: гаметофит прикрепляется к субстрату нитевидными ризоидами. Вода и минеральные соли поглощаются всей поверхностью тела, в том числе и ризоидами. А это значит, что ризоиды – не настоящие корни, они лишь закрепляют растение в грунте. (В настоящих корнях, как и в настоящих стеблях или листьях имеются проводящие ткани). Поэтому стебли и листья мохообразных не гомологичны стеблям и листьям сосудистых растений, у которых эти органы входят в состав диплоидного спорофита, а не гаплоидного гаметофита.
Поможем написать работу на аналогичную тему
5. Спорофит прикреплён к гаметофиту, полностью зависит от него и питается за его счёт
6. Споры на спорофите образуются в споровой коробочке, расположенной на конце тонкой ножки, возвышающейся над гаметофитом
7. Встречаются главным образом в сырых затенённых местах. Могут выдерживать периоды засухи, используя для этой цели какие-то не совсем понятные механизмы.
| Класс Hepaticae или Hepaticopsida (печеночные мхи) | Класс Musci или Bryopsida (лиственные мхи) |
| Гаметофит представлен уплощённым образованием, форма которого варьирует от талломной (очень редко) до «олиственной» со стеблем (у большинства видов); имеются переходные формы | Гаметофит «олиственный», имеет стебель и обычно гораздо более дифференцирован, чем у печеночных мхов |
| «Листья» расположены вдоль стебля тремя рядами | «Листья» расположены тесной спиралью |
| Ризоиды одноклеточные | Ризоиды многоклеточные |
| Для рассеивания спор коробочка спорофита раскрывается на четыре створки; рассеиванию спор помогают специальные пружинки - элатеры | Для рассеивания спор из коробочки спорофита выработался сложный механизм, который срабатывает в сухую погоду; функционирование этого механизма обеспечивается зубцами перистома и порами; элатеры отсутствуют |
| Примеры: Pellia – талломный печеночный мох Marchantia – талломный печеночный мох; антеридии и архегонии расположены на своеобразных выростах – подставках, возвышающихся над талломом Lophocolea – облиственный печеночный мох; обычно встречается на гниющей древесине | Funaria, Mnium – обыкновенный лесной мох, похожий по внешнему виду на фунарию Sphagnum – торфяной болотный мох; во влажной кислой среде (на болотах) образует слои торфа |
Чередование поколений
Листостебельное фотосинтезирующее тело мха представляет собой гаметофит. На нём развиваются антеридии (мужские половые органы) и архегонии (женские половые органы). В антеридиях образуются двужгутиковые сперматозоиды, в архегониях – крупные яйцеклетки. При помощи воды (в сырую погоду) сперматозоиды выходят из антеридиев. Один из них проникает в архегоний и оплодотворяет яйцеклетку. Из образовавшейся зиготы вырастает бесполое поколение – спорофит. Он представлен коробочкой на ножке, которая вырастает на гаметофите. Спорофит не способен к фотосинтезу и питается за счёт гаметофита. В коробочке образуются гаплоидные споры. При прорастании споры формируется протонема – тонкая разветвлённая нить, на которой образуются почки, дающие начало новым гаметофитам.
Цикл развития мха
отдел моховидные
Болота сосредоточены в основном в лесной зоне северного полушария, а также во влажных экваториальных районах Африки и Южной Америки. Общая площадь болот в мире – 350 млн га. На территории нашей страны сосредоточено около 3/4 мировых болот. Основной фон растительности верховых болот составляют сфагновые мхи.
В мире насчитывается 25 тысяч видов мохообразных, из них 15–20 тысяч видов листостебельных (зеленых и сфагновых) мхов. По количеству видов среди высших растений мхи занимают второе место после цветковых (покрытосеменных). Моховидные произошли 350 млн лет назад от первых наземных растений – псилофитов, потомков прибрежных водорослей. Это многолетние растения, обычно низкорослые, их размеры от 1 мм до нескольких см; они имеют стебель, листья, корней нет. Обитают во влажных местах.
Мхи отличаются простой внутренней организацией. В их теле можно обнаружить ассимиляционную (основную) ткань, содержащую хлоропласты, а также слабо выраженные проводящие, механические, запасающие и покровные ткани. Мхи прикрепляются к почве тонкими нитевидными одноклеточными или многоклеточными выростами, расположенными на нижней части стебля, ризоидами, через которые всасываются питательные вещества из почвы.
В отличие от настоящих корней многоклеточные ризоиды состоят из одинаковых клеток и лишены проводящих тканей.
-
Класс Hepatiopsida, особенности строения, размножения, цикла воспроизведения. Важнейшие представители.
Класс печеночники - hepaticopsida. Общее число видов около 10 000 (235 родов). Их подразделяют на 7 порядков. Важнейшие из них следующие: Калобриальные -- Calabryales, Юнгерманиальные -- Jungermanniales, Маршанциальные -- Marchantiales, Сферокарпальные -- Sphaerocarpales, Риччиаль-ные -- Ricctales.
По данным морфологического и ботанико-географического анализов, виды печеночников, равно как и антоцеротовых, очень древние. Здесь явно господствуют монотипные или олиготипные роды, а иногда и семейства Из числа немногих политипных таксонов печеночников необходимо отметить лишь некоторые семейства и роды юнгерманиальных. Полиморфизм юнгерманиальных свидетельствует об их относительной молодости и процветании. Значит, наряду с архаичными, реликтовыми, т. е. угасающими таксонами, представлены и развивающиеся.
В целом же юнгерманиальные подразделяют на 25 семейств, включающих 180 родов. Общее число видов превышает 8000, что составляет около 80 % всех печеночников.
По данным палеонтологии, наиболее примитивные (талломные) формы печеночников нужно отнести к девону. Более совершенные формы, близкие к маршанциальным, найдены в пермских отложениях. Типичные маршанциальные обнаружены в начале юрского периода. Юнгерманиальные--наиболее молодая группа; ископаемые остатки известны с конца мезозойской эры.
В географическом распространении печеночники практически не имеют границ. Класс в целом характеризуется как космополитический. При этом нужно отметить, что некоторые роды и виды имеют дизъюнктивное распространение, т. е. встречаются как бы разорвано -- на разных континентах: в Южной Америке, Австралии, Южной Африке, Юго-Восточной Индии.
Такой характер распространении неоспоримо свидетельствует о большой древности этих групп растений. На древность печеночников указывает и примитивная слоевищная структура тела и число хромосом (п = 4). Такой же набор хромосом имеют и некоторые зеленые водоросли. Однако слоевище у печеночников высоко специализировано.
В качестве представителя печеночников рассмотрим маршанцию. Как и некоторые другие печеночные мхи, маршанция -- исключительно слоевищное растение (рис. 3).
Вегетативное тело спинно-брюшной симметрии, пластинчатое, дихотомически ветвящееся, 10--12 см и длину. На морфологически верхней стороне таллома развиваются на ножках особые образования -- подставки, а на них -- органы полового размножения. На одних экземплярах подставки имеют форму девятилучевой звездочки, между лучами которой на нижней стороне расположены архегонии. На других экземплярах вырастают мужские подставки, имеющие форму восьмиугольных дисков или щитков. Антеридии погружены в литеридиальные полости на верхней стороне диска -- подставки.
Рис. 3. Цикл развития маршанции: 1 -- мужской гаметофит, 2--антеридий, 3 -- сперматозоид, 4 -- женский гаметофит, 5 -- архегоннй, 6 -- зигота, 7 -- начальная стадия развития спорогона, 8 -- спорогенная клетка, 9-- развитие из спорогенной клетки четырех спор путем мейоза, 12 -- раскрывшийся спороген, 13--прорастание споры, 14 -- молодой таллом
Макро- и микроскопическое строение пластинчатого таллома печеночников довольно сложное. С обеих сторон находится однослойная эпидерма, а внутри--крупноклеточная бесцветная паренхима и хлорофиллоносные клетки. Верхняя эпидерма несет вентиляционные отверстия-- устьица, образованные бесцветными клетками, расположенными друг над другом в четыре ряда. Под эпидермой находятся воздушные камеры. На дне этих камер развиваются нитевидные выросты-- ассимиляторы. Они состоят из хлорофиллоносных клеток. От нижней эпидермы отходят выросты, четко различающиеся между собой. Одни из них бесцветные одноклеточные ризоиды -- простые, и язычковые. Другие, более или менее окрашенные н(красноватые или зеленоватые),-- многоклеточные чешуйки, именуемые амфигастриями. Бриологи считают, что амфигастрии представляют собой видоизмененные и редуцированные «листья» -- филлоиды. Это дает основание полагать, что слоевищные печеночники (типа маршанции) произошли от « листостебельных».