ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 04.12.2023
Просмотров: 123
Скачиваний: 3
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
Затем в контакт со 2-м кодоном иРНК, сидящем на А-site, вступает та тРНК, антикодон которой комплементарен ему. Начинается элонгация, за которую отвечает энзим большой субъединицы – пептидилтрансфераза, основу которой составляет рРНК (поэтому-то он и называют - рибозим). С ее помощью α-аминогруппа новой аминоацил-тРНК в А-участке осуществляет нуклеофильную атаку этерифицированной карбоксильной группы первой аминоацил-тРНК, занимающей Р-site, образуется дипептид, связанный с тРНК2 и сидящий на аминоацильном локусе. Свободная тРНК1 покидает Р-участок.
Синтез:

Схема синтеза и созревания полипептида
Затем осуществляется транслокация, которая происходит с помощью фактора элонгации 2 (ЕF-2, транслоказа) и энергии гидролиза ГТФ, в результате происходит перемещение нити иРНК таким образом, что ее второй кодон оказывается на пептидильном участке вместе с дипептидом-тРНК, на А-site – третий триплет иРНК, способный к взаимодействию с соответствующей аминоацил-тРНК.
После многих подобных циклов элонгации и транслокации, в результате которых синтезируется полипептидная цепь, на А-локус садится нонсенс (терминирующий) кодон иРНК (УАА, УАГ или УГА), что останавливает ее рост. С помощью R-факторов (факторов терминации) гидролизуется связь между полипептидом и тРНК, занимающей Р-участок, а рибосома диссоциирует на свои субъединицы (Схема 4.4.1).
 В дальнейшем начинается созревание полипептидных нитей. В клетках эукариот многие белки синтезируются в виде предшественников, созревание которых представляет частичный гидролиз. Например:
В дальнейшем начинается созревание полипептидных нитей. В клетках эукариот многие белки синтезируются в виде предшественников, созревание которых представляет частичный гидролиз. Например:Некоторые пробелки подвергаются химической модификации: пролины проколлагена гидроксилируются, что обеспечивает зрелость коллагена. Известны и другие варианты посттрансляционных модификаций (гликозилирование, фосфорилирование, ацетилирование и т.д.), когда образуются различные простые и сложные протеины (приложение, Табл.11).
Параллельно происходит усложнение вторичной структуры, формирование (фолдинг) пространственной укладки с помощью шаперонов – белков, отвечающих за правильный характер сворачивания полипептидных цепей; кроме того эти протеины помогают вновь синтезированным нитям еще в развернутом состоянии преодолевать мембраны и попадать из цитозоля в органоиды.
Если почему-то нарушается структурирование полипептидных цепей
, то возникшие при этом белки не только могут выполнять свои функции, но и производят токсический эффект. Примером могут служить нейродегенеративные заболевания, в основе которых лежат преобразования α-спиралей прионов – белков нервной ткани в β-складчатую структуру, что придает данным протеинам инфекционные свойства. При этом высока вероятность особой роли в таких процессах соответствующих шаперонов.
Пусковым механизмом подобных болезней является неадекватное количество инфекционной формы приона. Это может быть вызвано:
а) мутацией его гена, что обычно носит спорадический характер (болезнь Крейтцфельдта-Якоба, синдром Герстманна-Штреусслера-Шейнкера);
б) проникновением инфекционной частицы извне алиментарным (ритуальный каннибализм провоцирует заражение болезнью Куру) или ятрогенным (нейрохирургические манипуляции, пересадка роговицы и других тканевых материалов от человека к человеку, терапия белковыми гормонами гипофиза) путями.
Поэтому прионные болезни и наследственные, и инфекционные одновременно.
4.5. Регуляция синтеза азотсодержащих биополимеров
Каждая из трех стадий этого сложного процесса имеет элементы, осуществляющие строгую регуляцию.
Первичным механизмом воздействия на скорость синтеза ДНК служит то, что репликация происходит в жестко определенное время в клетках, готовящихся к делению. В регуляции вступления в S-фазу участвуют циклические пуриновые нуклеотиды (ц-АМФ, ц-ГМФ), а также появление в этот момент большого количества ДНК-полимеразы α, усиление активности тиоредоксинредуктазной системы, участвующей в образовании субстратов генеза ДНК – дезоксирибонуклеозидтрифосфатов. При выходе из S-фазы деятельность этих энзимов резко снижается.
На стадии транскрипции для регуляции используются механизмы: а) изменения скорости экспрессии генов или инициация их репрессии; б) альтернативного сплайсинга. В первом случае исполнителями служат энхансеры (возбудители) и сайленсеры (замедлители). Экспрессия генов подразумевает его транскрипцию в иРНК с последующей трансляцией. Где и когда будет производиться этот процесс – определяют специфические некодирующие участки ДНК или особые белки, которые связываются с энхансерами, что стимулирует экспрессию. У каждого гена имеется по крайней мере один подобный возбудитель, который представляет фрагмент цепи ДНК, состоящий из нескольких сотен мононуклеотидов, может располагаться на различном расстоянии от регулируемого гена.
Гетерогенная ядерная ДНК – предшественница иРНК может включать до нескольких десятков интронов и экзонов. При взаимодействии этого соединения с малыми ядерными РНК, интроны вычленяются, а вторые сшиваются, причем последние могут соединяться в разных последовательностях, что приводит к образованию разных иРНК, кодирующих разные белки. Это наблюдаемое для многих генов явление получило название альтернативного сплайсинга, а количество зрелых иРНК, отличных друг от друга, может приближаться к величине 2n, где n – число экзонов. Случаи подобного синтеза наблюдаются в мышечной, нервной тканях, при образовании белков клеточной адгезии.
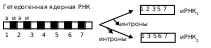
Нарушение же точности сплайсинга иРНК, вызванное мутацией, может препятствовать трансляции и созреванию белковых молекул.
Кроме того, существуют три основных способа регулирования этой стадии. Первый – за счет сродства иРНК к инициирующей рибосоме и факторам инициации. Второй – негативная регуляция с помощью белков – репрессоров, которые, связываясь с иРНК, блокируют инициацию (трансляционная репрессия). Третий механизм – тотальная регуляция синтеза всей совокупности иРНК клетки посредством модификации факторов инициации.
В первом случае, если сродство иРНК слабее, то способность инициации низкая, продукция белка невелика. Структурные же протеины мембран, требуемые в большом количестве, кодируются сильными иРНК, а специализированные ферменты и регуляторные белки – слабыми иРНК.
Механизм трансляционной репрессии заключается в том, что белок-репрессор специфически связывается с тем локусом иРНК, который взаимодействует обычно с рибосомой, тем самым мешает этому процессу и тормозит или блокирует полностью инициацию. Иногда в качестве репрессора выступает свой же продукт иРНК.
Наиболее обычный путь тотальной регуляции белкового синтеза у эукариот – это активация специальной фосфокиназы, которая фосфорилирует фактор инициации iF2. Последний в норме способствует связыванию аминоацил-тРНК с малой субъединицей рибосомы. Модификация фактора мешает возможности начала трансляции. Сигналами для активации фосфокиназы в клетке являются недостаток ростовых факторов, аминокислотное голодание, дефицит железа, вирусные инфекции, тепловой шок и другие виды стрессовых воздействий.
За последние годы исследователям открылась масса новых фактов, свидетельствующих о решающем вкладе не генной активности, а трансляционной регуляции, в частности под влиянием различных гормонов, при старении, при смене фаз клеточного цикла, в процессах развития и клеточной дифференцировки.
4.6. Причины нарушений генеза азотсодержащих биополимеров
Основными субстратами и в синтезе белков, и в образовании нуклеотидов являются аминокислоты, следовательно, одной из причин, провоцирующих снижение скорости вышеназванных процессов, будет недостаточное поступление или нарушение соотношения аминокислот в пище (белковое или полное голодание) или дисфункции ЖКТ [снижение секреции пищеварительных соков (желудочного, поджелудочного, кишечного) или нарушение целостности слизистой тонкой кишки], в результате чего угнетается усвояемость и всасывание этих соединений.
Спровоцировать изменения в течении синтеза данных биополимеров могут вещества, обладающие способностью воздействия на деятельность ферментов – участников этих процессов. Ими могут быть гормоны (СТГ, инсулин и др.) или лекарства (антибиотики, например, тетрациклин нарушает связывание аминоацил-тРНК с рибосомой, эритромицин тормозит образование пептидных связей), токсины (дифтерийный токсин тормозит транслокацию и пептидная цепь остается в А-site рибосомы). Выраженность этих, как известно, определяется дозой.
Но особое место среди причин занимает так называемые мутагены – факторы, приводящие к повреждениям первичной структуры ДНК. Их природа различна. Можно выделить физические (чрезмерно высокая или низкая температура, радиация, ультразвук, рентгеновское, инфракрасное, ультрафиолетовое, α-, γ- и другие виды излучений); химические (нитраты, нитриты, анилиновые красители, продукты переработки нефти и другие углеводороды, в т.ч. ароматические, соли тяжелых металлов, свободные радикалы, лекарства (иммунодепрессанты, цитостатики), пестициды, гербициды и т.д.); биологические (вирусы краснухи, гриппа, кори; антигены – токсины бактерий, простейших) факторы.
Последствия их воздействий различны по типам наследования молекулярных изменений, по влиянию на организм (см. «Патохимия наследственных болезней», с.9).
В настоящее время стали обращать внимание не только на характер нарушений в нуклеотидной последовательности ДНК, но и на способность к восстановлению поврежденных участков (репарацию). Оказалось, что иногда в основе развития заболевания лежит как раз невозможность организма устранить возникшие аномалии в цепи ДНК. Примерами таких недугов являются болезни преждевременного старения (прогерия), пигментная ксеродермия и т.д. (см. «Патохимия наследственных болезней», с.16).
4.7. Принципы профилактики и терапии наследственных болезней
Как отмечено выше, немалую роль в развитии патологических процессов, играют мутации, которые в зависимости от протяженности повреждений в ДНК, делят на геномные, хромосомные, генные. Последние способны передаваться по наследству, другие же из-за грубых аномалий, в первую очередь, в половой сфере, - не наследуются. Дефекты генов могут быть одиночные (моногенные) или множественные (полигенные). Первые и провоцируют наследственные заболевания, для вторых характерна лишь наследственная предрасположенность (атеросклероз, сахарный диабет, гипертоническая болезнь, варианты злокачественных новообразований и т.д.). Отсюда и принципы их профилактики и лечения различны.