ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 04.12.2023
Просмотров: 126
Скачиваний: 3
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
а) первичную подагру - наследственное заболевание, известное под названием ювенильная гиперурикемия, или синдром Lesch – Nyhan (учебное пособие «Питохимия наследственных болезней», с.71) у таких младенцев блокирована гипоксантингуанинфосфорибозилтрансфераза, отвечающая за использование пуринов освободившихся при распаде тканевых полинуклеотидов. Нарушение включения этих соединений в новые мононуклеотиды заставляет их преобразовываться в мочевую кислоту.
б) Вторичную подагру провоцируют погрешности в питании (избыток мяса, печени, почек, креветок, моллюсков, икры и молоки рыб и т.д.), усиленный распад тканей (при далеко зашедших злокачественных новообразованиях, лейкозах, их лечении цитостатиками, радиотерапией, или тяжелых инфекциях, ожоговой болезни), нарушение экскреторной функции почек и т.д. В местностях с избытком Мо2+ в пищевых продуктах возможно развитие молибденовой подагры (схема 3.5.1., д) вследствие активации ксантин ДГ.
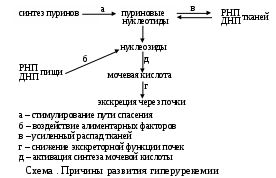
Возникающая при вышеописанных обстоятельствах гиперурикемия грозит следующими последствиями. Растворимость уратов в плазме крови ограничена, увеличение их концентрации свыше 0,5 ммоль/л способствует осаждению, кристаллизации. Наиболее благоприятным для этого местами будут участки, где замедлена скорость кровотока (суставы стоп)); кроме того, мочевая кислота, имея в составе гидроксильные группы, легко образует водородные связи с соединениями, также включающими подобные полярные группы, в том числе с гетерополисахаридами соединительно-тканных образований суставов, задерживаясь в них. Кристаллы уратов раздражают нервные рецепторы, вызывают боль, красноту, отек и другие симптомы воспаления. Покраснение является следствием расширения сосудов, что уменьшает кровоток, способствует гипоксии, накоплению недоокисленных продуктов (в первую очередь - лактата). Возникающее подкисление среды нарушает и так слабую растворимость уратов, создавая порочный круг.
Эти соли откладываются не только в мелких суставах, но и в коже (ушей), локтевых, коленных сумках, сухожилиях, образуя специфические узелки (tophi urici), что провоцирует приступы боли; избыточная секреция их почками способствует развитию нефропатии вплоть до мочекаменной болезни.
Противопололжный симптом - гипоурикемии - встречается очень редко, обычно провоцируется снижением активности ксантинДГ или повреждением печени; протекает бессимптомно, интеллект не нарушается.
Кроме сцепленной с Х – хромосомой ювенильной гиперурикемии, описаны еще следующие наследственные заболевания пуринового обмена: недостаточность аденозиндезаминазы, ксантинурия (учебное пособие «Патохимия наследственных болезней», С. 71).
Что касается обмена пиримидинов, то в настоящее время описана лишь оротацидурия (учебное пособие «Патохимия наследственных болезней», с.73).
Тесты к главе 3
-
Что такое цитидин?
а) сложный эфир
б) простой эфир
в) мононуклеотид
*г) нуклеозид
д) азотистое основание.
2. Если гуанин образовал N-гликозидную связь с дезоксирибозой, как называется продукт?
а) ГМФ
б) дГМФ
в) гуанин
г) гуанидин
*д) д-гуанозин
3. Укажите динуклеотиды:
*А) НАД+
*б) ФАД
в) ТДФ
*г) НS-КоА
д) АТФ
4. С помощью какой связи образуется динуклеотид, не способный к полимеризации?
а) сложноэфирной
*б) ангидридной
в) амидной
г) водородной
д) нуклеотидной
5. Укажите основную роль динуклеотидов.
а) гормоны
*б) коферменты
в) холоферменты
г) макроэрги
д) субстраты в синтезе полипептидов
6. Перечислите все возможные связи в нуклеиновых кислотах.
а) О-гликозидная
*б) N-гликозидная
*в) ионная
*г) водородная
*д) фосфодиэфирная
7. С разрыва какой связи начинается распад пищевых нуклеопротеидов в ЖКТ?
а) N-гликозидный
б) О-гликозидной
*в) ионной
г) амидной
д) фосфодиэфирной.
8. В каком органе ЖКТ образуется эндонуклеазы?
а) в желудке
б) в печени
*в) в поджелудочной железе
г) в слюнных железах
д) в железах тонкого кишечника.
9. Укажите основной продукт гидролиза нуклеиновых кислот в ЖКТ.
а) динуклеотид
б) мононуклеотид
*в) нуклеозид
г) аминокислота
д) пурины
10. Работой какого фермента отличается гидролиз тканевых полинуклеотидов от их пищевых аналогов?
а) РНК-аза
б) ДНКаза
в) дезамидимаза
*г) 5’ – нуклеотидаза
д) кислая фосфатаза
11. Укажите компоненты ДНК:
*а) ТМФ
б) АМФ
в) УМФ
*г) дГМФ
*д) дЦМФ
12. Как называются информативные участки генов?
а) сайленсеры
б) интроны
в) энхансеры
*г) экзоны
д) полиндромы
13. Укажите верные утверждения:
а) дигидроурацил - минорное основание, входящее в состав мононуклеотидов м-ДНК.
*б) дигидроурацил - минорное основание, входящее в состав мононуклеотидов т-РНК.
*в) т–РНК – долгожитель среди всех РНК;
*г) т–РНК содержатся и в цитоплазме, и в митохондриях;
д) концентрация м–ДНК изменяется с возрастом.
14. Что такое антикодон?
а) трипептид
*б) тринуклеотид
в) триплет рРНК, комплементарный кодогену
*г) триплет тРНК, комплементарный смысловому кодону и РНК
д) триплет гена
15. Что защищает иРНК от гидролиза нуклеазами?
а) нетранслирующие последовательности
б) терминирующий кодон
*в) полиаденилат
г) энхансер
д) транскриптон
16. Что такое информосома?
а) комплекс иРНК с рРНК
б) комплекс иРНК с тРНК
в) комплекс иРНК с транскриптоном
*г) комплекс иРНК с протеинами
д) комплекс рРНК с протеинами.
17. Молярная доля пуринов в нитях ДНК равна молярной доле пиримидинов. Кем предложен этот постулат?
а) Уотсон
б) Крик
в) Белозерский
г) Кребс
*д) Чаргафф
18. Какой вид имеет третичная структура мДНК?
а) двойная спираль
б) суперспираль
в) клеверный лист
г) триплекс
*д) кольцевой дуплекс
19. Как называется участок я-ДНК, располагающийся между нуклеосомами?
*а) линкер
б) ню-частица
в) энхансер
г) интрон
д) октамер
20. Укажите физико-химические свойства ДНК.
*а) хорошая растворимость в воде
б) плохая растворимость в воде
*в) высокая плотность
г) денатурация при температуре около 500
*д) денатурация при температуре выше 800
21. Что такое рестрикция?
а) способность ДНК к самообновлению
б) способность РНК к самообновлению
в) синтез ДНК на РНК
г) способность РНК к синтезу минорных оснований
*д) способность с помощью нуклеаз вычленять из молекулы ДНК поврежденные участки
22. Катион какого металла активирует ксантинДГ?
а) Мn
б) Мg
*в) Мо
г) Fе
д) Си
23. С какой реакции начинается преобразование аденозина в мочевую кислоту?
а) с восстановительного дезаминирования
б) с окислительного дезаминирования
*в) с гидролитического дезаминирования
г) с гидролиза N-гликозидной связи
д) с гидролиза О-гликозидной связи.
24. Какой фермент катализирует реакцию высвобождения урацила из уридина?
а) дигидроурацилДГ
б) дигидропиримидиназа
в) аминотрансфераза
г) ксантинДГ
*д) нуклеозидфосфорилаза
25. Укажите конечные продукты распада пиримидинов:
*а) соли аммония
*б) гидрокарбонаты
*в) мочевина
г) ураты
*д) вода
26. Укажите нормальное содержание мочевой кислоты в плазме крови:
а) до 5 мг%
б) до 5ммоль/л
в) до 0,5 мг%
*г) до 0,5 ммоль/л
д) до 0,5 мкмоль/л
27. Назовите термин, обозначающий повышенное содержание уратов в плазме крови:
а) гиперкарбамидемия
*б) гиперурикемия
в) гиперазотемия
г) гипоуринемия
д) гипоазотемия
28. Что может служить источником рибозы при синтезе мононуклеотидов?
*а) пищевые углеводы
б) гликолиз
в) ЦТК
*г) ПФП
д) фосфорилирование дезоксирибозы
29. Укажите аминокислоты, не используемые в синтезе пиримидинов.
*а) гли
*б) ала
в) асп
г) глн
д) глу
30. Укажите аминокислоты, атомы которых используются в синтезе пуринов:
а) ала
*б) асп
*в) глн
*г) гли
д) арг
31. В каких процессах участвует карбамоилфосфатсинтаза?
а) в синтезе пуринов
б) в синтезе пиримидинов
в) в синтезе ВЖК
*г) в синтезе мочевины
д) в синтезе аминосахаров
32. Звеном какой системы служит тиоредоксинредуктаза?
а) синтеза пуринов
б) синтеза пиримидинов
в) синтеза нуклеозидтрифосфатов
*г) преобразования рибонуклеотидов в дезоксирибибонуклеотиды
д) распад дезоксирибонуклеозидфосфатов
33. Активная форма какого витамина участвует в синтезе ТМФ?
а) С
*б) Вс (фолиевой кислоты)
в) тиамина (В1)
г) селена
д) РР (никотинамида)
34. Укажите болезни, в основе которых лежат нарушения в метаболизме пуринов:
*а) синдром Lasch – Nyhanc
б) синдром Хюрлера
*в) подагра
*г) ксантиурия
д) оротацидурия
35. Укажите характерные признаки первичной подагры:
*а) гиперурикемия
б) гипоурикемия
*в) очень низкий интеллект
г) интеллект не страдает
д) аутоагрессивность
36. Укажите причины вторичной подагры:
а) блок гипоксантингуанинфосфорибозилтрансферазы
*б) излишества морских продуктов, печени, почек, алкоголя в питании
*в) IV стадия злокачественного образования
*г) нарушение фильтрационной способности почек
д) угнетение синтеза мочевой кислоты
37. Генетический дефект какого фермента обуславливает содержание младенцев в стерильных условиях (специальных скафандрах)?
а) дигидрониримидиназа
*б) аденозиндезаминаза
в) ксантинДГ
г) гипоксантингуанинфосфорибозилтрансферазы
д) аденозинфосфорибозилтрансфераза
38. Блок каких ферментов лежит в основе развития оросацидурии?
а) дигидрооротатДГ
*б) оротатфосфорибозилтрансфераза
в) дигидропиримидиназа
*г) ОМФ – декарбоксилаза
д) нуклеозидфосфорилаза.
Глава 4. Синтез азотсодержащих биополимеров
Схематически все процессы образования полипептидов и полинуклеотидов укладываются в следующую схему:
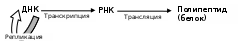
- где над стрелками обозначены названия соответствующих стадий.
Общие принципы реакций
Все биополимеры (полипептиды, гетерополисахариды, полинуклеотиды) имеют организацию, - специфическую упаковку в пространстве, поэтому их синтез включает дополнительно и стадию структурирования. А если их звенья еще и отличаются друг от друга, отсюда, чтобы вновь образующийся полимер включал их в строго определенном порядке, необходима матрица, что и отличает генез белков и нуклеиновых кислот от других подобных процессов. Кроме того, любой синтез требует затрат энергии, причем часто в этих случаях используются не обычные макроэрги, а происходит с их помощью предварительная активация субстратов. Исходя из этих предпосылок, можно выделить следующие общие принципы, характерные для этих процессов:
-
Наличие матрицы: в синтезе дочерней ДНК ею служит вся материнская ДНК; для образования РНК используется фрагмент ДНК (ген, или транскриптон); аминокислотная последовательность определяется набором триплетов иРНК.
-
Субстраты предварительно активируются: на первой стадии используются дезоксирибонуклеотидтрифосфаты (дАТФ, дГТФ, дЦТФ, ТТФ), генез РНК требует наличия соответственно АТФ, ГТФ, ЦТФ, УТФ, полипептиды получаются из аминоацил-тРНК.
-
Основные ферменты, обеспечивающие собственно полимеризацию, относятся к классу трансфераз и называются ДНК-, РНК-полимеразы и пептидилтрансфераза.
-
Сложность образования биополимера выделять в этом процессе две фазы: собственно синтез (как бы начерно) и созревание – укладку в пространстве, химическую модификацию.
-
В собственно синтезе выделяют следующие стадии: а) инициацию – начало; б) элонгацию (longus - длинный) – наращивание цепи; в) терминацию – остановку реакций.
Репликация ДНК
Репликация – синтез дочерней ДНК с использованием в качестве матрицы всей молекулы материнской ДНК. С этого процесса и начинается деление клетки в S-фазу (синтетическую). В основе репликации лежат три принципа: а) комплементарности – азотистые основания образующейся дочерней цепи должны обладать химическим и геометрическим соответствием с подобными веществами материнской нити; полуконсервативности – каждая из синтезированных молекул ДНК состоит из одной материнской и одной дочерней; в) однонаправленности – считывание матрицы идет от 5' к 3' концу.
Чтобы порядок азотистых оснований, находящихся внутри нитей ДНК, можно было считывать, необходимо разорвать водородные связи, соединяющие между собой цепи и лишить их спирализации. Для этих целей используются следующие ферменты: топоизомераза и хеликаза. Первая гидролизует на одном из участков полинуклеотида фосфодиэфирная связь, что позволяет этому фрагменту раскрутиться относительно параллельно лежащего:
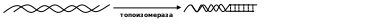
а затем восстановить связь. Хеликаза разрушает водородные взаимодействия между противолежащими основаниями участков двух цепей:
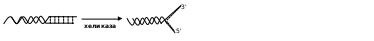
Под ее действием фрагменты расходятся, образуя репликативную вилку. Затем с помощью праймазы (primer - затравка) синтезируется олигорибонуклеотид – небольшое соединение, состоящее (обратите внимание) из 8-10 монорибонуклеотидов. Оно служит местом, куда крепится основной фермент – ДНК-полимераза. Из-за антипараллельности цепей данные структуры будут локализоваться в разных местах: одна у конца, другая несколько в глубине противоположной нити:
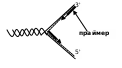
Все дело в том, что ДНК-полимеразы не могут начинать синтез, а способны только добавлять дезоксирибонуклеотидные звенья к 3'-концу уже имеющейся цепи, чем и является праймер. Описано 3 класса данных энзимов: в ядрах присутствуют полимеразы альфа (Роl α), ответственные за хромосомную репликацию, полимеразы бета (Роl β), которые используются при необходимости при репарации. Третий их представитель – ДНК-полимераза гамма (Роl γ) осуществляет синтез кольцевого генома митохондрий.
Присоединение каждого нового нуклеотидного остатка к 3'-концу растущей цепи сопровождается гидролизом макроэргической связи в дНТФ и отщеплением пирофосфата.
Важная деталь – генетический материал живых организмов имеет огромные размеры, но реплицируется с высокой точностью. В среднем при воспроизведении генома (ДНК длиной более 3 млрд пар нуклеотидов) возникает не более 3-х ошибок благодаря наличию специальных механизмов, осуществляющих необходимую коррекцию. Суть последней в том, что ДНК-полимеразы дважды проверяют соответствие каждого нуклеотида матрице: один раз перед включением его в состав растущей цепи и второй – перед присоединением нового нуклеотида. Очередная фосфодиэфирная связь синтезируется лишь в том случае, если последний мономер удлиняющейся дочерней нити ДНК образовал правильную (комплементарную) пару с соответствующим нуклеотидом матрицы. Если же на предыдущей стадии произошло ошибочное спаривание оснований, то фермент возвращает последнее добавленное звено, после чего освободившееся место занимает правильный нуклеотид.
Параллельно с ростом цепи продолжается деспирализация и разделение цепей с помощью топоизомеразы и хеликазы, удлинение репликативной вилки. А так как процесс осуществляется лишь в одном (5' – 3') направлении, что позволяет происходить этому явлению непрерывно только на одной из матричных нитей. На антипараллельной синтез ограничивается сравнительно короткими фрагментами (100-1000 мононуклеотидов), названными по имени обнаружившего их ученого фрагментами Оказаки (с постоянным предварительным синтезом праймеров):
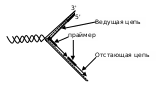
Так продолжается до тех пор, пока вся матрица не выполнит свою функцию, причем во время репликации специальные нуклеазы вычленяют праймеры. Если позволяет направление, ДНК-полимеразы застраивают возникшие бреши, а сшивание сближенных фрагментов обеспечивается ДНК-лигазами – так завершается собственно синтез (стадия терминации).
В тех концах дочерних цепей, где находились праймеры, которые позднее были удалены, не происходит достраивания дезоксирибонуклеотидами (невозможно считывание в обратном направлении), отсюда при каждом делении клетки молекулы новых цепей укорачиваются на 10-20 нуклеотидов, но объем информации при этом не уменьшается, так как потерянные участки ее не несли. Дело в том, что когда в эмбриональных тканях шли усиленные митозы параллельно с этим осуществлялись обратные транскрипции. С помощью теломеразы концы ДНК наращивались теломерными повторами ТТАGGG, роль матрицы при этом выполняла РНК, включающая 450 мононуклеотидов. Поскольку многократно повторенные теломерные последовательности не являются кодирующими, их утрата в процессе деления не приводит к потере информативных участков, тем самым обеспечивается передача генетического материала в поколениях клеток.
Однако в родившемся организме активность теломераз регистрируется лишь в половых, раковых, стволовых клетках, в соматических же эти ферменты не работают. Теломеры постепенно укорачиваются, что в конце концов запускает процессы остановки клеточного цикла и провоцирует апоптоз. Интересный факт: ученые, исследующие рак, до сих пор используют культуру клеток Неlа из опухоли женщины по имени Генриетта Лакс. Эта больная умерла в 1951 году, но клетки ее новообразования продолжают делиться и собираются жить вечно.
Происходящее в момент деления клетки удлинение синтезирующихся нитей ДНК сопровождается их созреванием, что включает спирализацию, суперспирализацию (закручивание цепей вокруг гистоновых нуклеосом), химическую модификацию (гидроксилирование, метилирование, гидрирование азотистых оснований) с образованием минорных оснований, соединение с помощью ионных связей с белками, катионами металлов (К+, Са++, Мg++, Мn++, Fе++, Сu++ и т.д.), которые стабилизируют или, наоборот, дестабилизируют вновь синтезированную молекулу.
Синтез и процессинг РНК
В отличии от ДНК, синтез которой происходит в момент деления клетки и служит источником генетической информации для последующих поколений, различные РНК необходимы для генерирования молекул белков. Процесс синтеза рибонуклеиновых кислот с использованием в качестве матрицы фрагментов ДНК называют транскрипцией. Все гены в хромосоме (ДНК с белками) в отсутствии факторов транскрипции находится в выключенном (репрессированном) состоянии, потому что нуклеосомы блокируют области инициирования каждого промотора. Ген будет транскрибироваться после вытеснения нуклеосомы и связывания. Та из двух цепей ДНК, на которой пойдет этот процесс, называется кодирующей. Инициация включает раскручивание участка, разрыв водородных связей, что раскрывает азотистые основания транскриптона, в котором выделяют промотор, собственно ген (единицу транскрипции), терминатор. Первая функциональная единица (промотор) служит для с ним РНК-полимеразы, которая объединяет рибонуклеотиды в последовательность, комплементарную кодирующей цепи гена. Момент терминации распознается терминирующим белком – р-фактором.
Интересно, что в районе промотора расположены сигнальные последовательности двух типов. Одна из них указывает, где должна начаться транскрипция, а другая определяет, как часто должно происходить это событие. Первый локус называют ТАТА-бокс, второй – СААТ-бокс. Кроме них регуляторная зона включает энхансеры или сайленсеры, изменяющие скорость транскрипции, а также гормончувствительные элементы (ГЧЭ), после взаимодействия с которыми кортикостероиды, андрогены, эстрогены, Т3, сАМФ регулируют экспрессию генов.
Практически все первичные транскрипты РНК у эукариот подвергаются сложному процесссингу, т.е. созреванию, начинающемуся в ядре. Будущая иРНК получается после копирования, реакций расщепления, вычленения, легирования, включения дополнительных адениловых нуклеотидов. Ведь свежеполученная РНК содержит неинформативные вставки, считанные с интронов генов, т.е. представляют чередование траслируемых и нетранслируемых участков (гетерогенная ядерная РНК). Процесс удаления интронов и последующего сшивания экзонов получил название сплайсинг (от англ. to splice – соединять концы, сшивать). Он требует наличия специальных ферментов, поэтому осуществляется в крупном рибонуклеопротеидном комплексе – тельце, содержащем РНК и белки, необходимые для эффективного сплайсинга. Эта структура называется сплайсосома. Кроме того к 5'-концу г/яРНК присоединяется 7-метилгуанозинфосфат (кэпирование), который позднее служит стартовой точкой синтеза белка, к 3'-концу надстраивается полиаденилатный хвост (около 200 мононуклеотидов), защищающий иРНК от действия нуклеаз. Параллельно иРНК уже вышедшая из ядра, плотно упаковывается в пространстве.
Первичный транскрипт рРНК не содержит интронов, интенсивно метилируется и расщепляется в ядрышке специфическими РНК-азами на 5S, 18S, 28-рРНК, затем они связываются с белками, образуя рибосомы. Подобным образом (путем частичного гидролиза) осуществляется из первичных транскриптов образование тРНК, формирование вторичной, третичной структуры, химическая модификация. Критическое значение имеет этап сплайсинга в области антикодона, т.к. от него зависит точность выполнения адапторной функции при синтезе белка.
4.4. Синтез полипептидов
Образно говоря, для осуществления двух предыдущих стадий использовался один язык – нуклеотидный, третья же стадия – синтез полипептидов – говорит на другом языке – языке аминокислот. Следовательно, требуется перевод. Поэтому данный процесс и называется трансляцией. Это строительство аминокислотной последовательности на иРНК в качестве матрицы.
Для успешного перевода сведений, записанных в генах ДНК, в строго специфический порядок аминокислот в белке используется биологический (генетический) код.
Положения генетического кода
-
Триплетность. В полипептидах всего 4 разных азотистых основания, аминокислот не менее 20, следовательно – один мононуклеотид не может нести информацию об одной аминокислоте; два (42=16) – тоже, и только количество тринуклеотидов (43=64) может соответствовать числу аминокислот. Причем 3 из них (УАА, УАГ, УГА) бессмысленные (стоп-сигналы) – не несут сведения ни о какой аминокислоте, а 61 – смысловой триплет.
-
Вырожденность. Информация об одной аминокислоте может быть заложена в 1-3, до 5 различных кодонах.
-
Однозначность. С одного кодона можно считать сведения только об одной аминокислоте.
-
Однонаправленность. Считывание производится только в одном направлении с 5'-конца к 3'-концу.
-
Код не перекрывающийся – один и тот же мононуклеотид не может входить в состав двух разных, рядом лежащих кодонов.
-
Код «без запятых» – два рядом расположенных триплета не отделяются друг от друга никакими знаками препинания.
-
Код не универсален, а альтернативен – триплеты ядерной ДНК, несущие сведения об определенных аминокислотах могут отличаться от соответствующих кодонов м-ДНК (см. «Патохимию наследственных болезней», с.18).
Обязательными участниками трансляции служат иРНК, рибосомы, тРНК и аминокислоты. Последние, чтобы стать настоящими субстратами, должны не только активироваться, но и связаться с той тРНК, антикодон которой несет информацию о ней.
Поэтому в клетке присутствуют специальные ферменты, осуществляющие этот процесс и каждый из которых обладает суперспецифичностью, механизм которой называют вторым генетическим кодом. Проверка правильности образования комплекса осуществляется на обеих стадиях, и если выясняется ошибочность, тут же происходит распад образовавшейся структуры:
Е + а/к + АТФ ↔ Е▪аминоацил
АМФ + ФФ
Е▪аминоацил
АМФ + тРНК ↔ Е + АМФ + аминоацил
тРНК,
где Е – фермент (аминоацил-тРНК-синтетаза), а/к – аминокислота.
Подготовленные таким способом аминокислоты подтягиваются к рибосомам, куда подходит и синтезированная в ядре иРНК. Когда рибосома не транслирует, она находится в диссоциированном состоянии, т.е. распадается на две неравные субчастицы: у эукариот их обозначают:
80S → 60S + 40S.
(S-единицы Svedberge, в них рассчитывается скорость седиментации – осаждения при ультрацентрифугировании)
Малая субъединица (40S) имеет два специальных локуса: аминоацильный (А) и пептидильный (Р). Инициация (Схема 4.4.1) начинается с того, что к этой структуре подходит иРНК и своим кэп-участком так крепится к ней, что ее первый оказывается на Р-, а второй на А-локусе. Этот процесс осуществляется при обязательном участии факторов инициации (iF-1 – iF-3).
У иРНК первым (инициирующим) кодоном всегда является триплет, несущий информацию о метионине (АУГ), отсюда из всех аминоацил-тРНК к пептидильному участку может подойти с последующим образованием водородных связей с этим кодоном только та, антикодон которой ему комплементарен. В итоге образуется комплекс: инициирующий тринуклеотид иРНК - аминоацил
тРНК – малая субъединица. Инициация завершается тем, что к нему присоединяется 60S субъединица, предотвращающая обратимость процесса.
Глава 4. Синтез азотсодержащих биополимеров
Схематически все процессы образования полипептидов и полинуклеотидов укладываются в следующую схему:
- где над стрелками обозначены названия соответствующих стадий.
Общие принципы реакций
Все биополимеры (полипептиды, гетерополисахариды, полинуклеотиды) имеют организацию, - специфическую упаковку в пространстве, поэтому их синтез включает дополнительно и стадию структурирования. А если их звенья еще и отличаются друг от друга, отсюда, чтобы вновь образующийся полимер включал их в строго определенном порядке, необходима матрица, что и отличает генез белков и нуклеиновых кислот от других подобных процессов. Кроме того, любой синтез требует затрат энергии, причем часто в этих случаях используются не обычные макроэрги, а происходит с их помощью предварительная активация субстратов. Исходя из этих предпосылок, можно выделить следующие общие принципы, характерные для этих процессов:
-
Наличие матрицы: в синтезе дочерней ДНК ею служит вся материнская ДНК; для образования РНК используется фрагмент ДНК (ген, или транскриптон); аминокислотная последовательность определяется набором триплетов иРНК.
-
Субстраты предварительно активируются: на первой стадии используются дезоксирибонуклеотидтрифосфаты (дАТФ, дГТФ, дЦТФ, ТТФ), генез РНК требует наличия соответственно АТФ, ГТФ, ЦТФ, УТФ, полипептиды получаются из аминоацил-тРНК.
-
Основные ферменты, обеспечивающие собственно полимеризацию, относятся к классу трансфераз и называются ДНК-, РНК-полимеразы и пептидилтрансфераза.
-
Сложность образования биополимера выделять в этом процессе две фазы: собственно синтез (как бы начерно) и созревание – укладку в пространстве, химическую модификацию.
-
В собственно синтезе выделяют следующие стадии: а) инициацию – начало; б) элонгацию (longus - длинный) – наращивание цепи; в) терминацию – остановку реакций.
Репликация ДНК
Репликация – синтез дочерней ДНК с использованием в качестве матрицы всей молекулы материнской ДНК. С этого процесса и начинается деление клетки в S-фазу (синтетическую). В основе репликации лежат три принципа: а) комплементарности – азотистые основания образующейся дочерней цепи должны обладать химическим и геометрическим соответствием с подобными веществами материнской нити; полуконсервативности – каждая из синтезированных молекул ДНК состоит из одной материнской и одной дочерней; в) однонаправленности – считывание матрицы идет от 5' к 3' концу.
Чтобы порядок азотистых оснований, находящихся внутри нитей ДНК, можно было считывать, необходимо разорвать водородные связи, соединяющие между собой цепи и лишить их спирализации. Для этих целей используются следующие ферменты: топоизомераза и хеликаза. Первая гидролизует на одном из участков полинуклеотида фосфодиэфирная связь, что позволяет этому фрагменту раскрутиться относительно параллельно лежащего:
а затем восстановить связь. Хеликаза разрушает водородные взаимодействия между противолежащими основаниями участков двух цепей:
Под ее действием фрагменты расходятся, образуя репликативную вилку. Затем с помощью праймазы (primer - затравка) синтезируется олигорибонуклеотид – небольшое соединение, состоящее (обратите внимание) из 8-10 монорибонуклеотидов. Оно служит местом, куда крепится основной фермент – ДНК-полимераза. Из-за антипараллельности цепей данные структуры будут локализоваться в разных местах: одна у конца, другая несколько в глубине противоположной нити:
Все дело в том, что ДНК-полимеразы не могут начинать синтез, а способны только добавлять дезоксирибонуклеотидные звенья к 3'-концу уже имеющейся цепи, чем и является праймер. Описано 3 класса данных энзимов: в ядрах присутствуют полимеразы альфа (Роl α), ответственные за хромосомную репликацию, полимеразы бета (Роl β), которые используются при необходимости при репарации. Третий их представитель – ДНК-полимераза гамма (Роl γ) осуществляет синтез кольцевого генома митохондрий.
Присоединение каждого нового нуклеотидного остатка к 3'-концу растущей цепи сопровождается гидролизом макроэргической связи в дНТФ и отщеплением пирофосфата.
Важная деталь – генетический материал живых организмов имеет огромные размеры, но реплицируется с высокой точностью. В среднем при воспроизведении генома (ДНК длиной более 3 млрд пар нуклеотидов) возникает не более 3-х ошибок благодаря наличию специальных механизмов, осуществляющих необходимую коррекцию. Суть последней в том, что ДНК-полимеразы дважды проверяют соответствие каждого нуклеотида матрице: один раз перед включением его в состав растущей цепи и второй – перед присоединением нового нуклеотида. Очередная фосфодиэфирная связь синтезируется лишь в том случае, если последний мономер удлиняющейся дочерней нити ДНК образовал правильную (комплементарную) пару с соответствующим нуклеотидом матрицы. Если же на предыдущей стадии произошло ошибочное спаривание оснований, то фермент возвращает последнее добавленное звено, после чего освободившееся место занимает правильный нуклеотид.
Параллельно с ростом цепи продолжается деспирализация и разделение цепей с помощью топоизомеразы и хеликазы, удлинение репликативной вилки. А так как процесс осуществляется лишь в одном (5' – 3') направлении, что позволяет происходить этому явлению непрерывно только на одной из матричных нитей. На антипараллельной синтез ограничивается сравнительно короткими фрагментами (100-1000 мононуклеотидов), названными по имени обнаружившего их ученого фрагментами Оказаки (с постоянным предварительным синтезом праймеров):
Так продолжается до тех пор, пока вся матрица не выполнит свою функцию, причем во время репликации специальные нуклеазы вычленяют праймеры. Если позволяет направление, ДНК-полимеразы застраивают возникшие бреши, а сшивание сближенных фрагментов обеспечивается ДНК-лигазами – так завершается собственно синтез (стадия терминации).
В тех концах дочерних цепей, где находились праймеры, которые позднее были удалены, не происходит достраивания дезоксирибонуклеотидами (невозможно считывание в обратном направлении), отсюда при каждом делении клетки молекулы новых цепей укорачиваются на 10-20 нуклеотидов, но объем информации при этом не уменьшается, так как потерянные участки ее не несли. Дело в том, что когда в эмбриональных тканях шли усиленные митозы параллельно с этим осуществлялись обратные транскрипции. С помощью теломеразы концы ДНК наращивались теломерными повторами ТТАGGG, роль матрицы при этом выполняла РНК, включающая 450 мононуклеотидов. Поскольку многократно повторенные теломерные последовательности не являются кодирующими, их утрата в процессе деления не приводит к потере информативных участков, тем самым обеспечивается передача генетического материала в поколениях клеток.
Однако в родившемся организме активность теломераз регистрируется лишь в половых, раковых, стволовых клетках, в соматических же эти ферменты не работают. Теломеры постепенно укорачиваются, что в конце концов запускает процессы остановки клеточного цикла и провоцирует апоптоз. Интересный факт: ученые, исследующие рак, до сих пор используют культуру клеток Неlа из опухоли женщины по имени Генриетта Лакс. Эта больная умерла в 1951 году, но клетки ее новообразования продолжают делиться и собираются жить вечно.
Происходящее в момент деления клетки удлинение синтезирующихся нитей ДНК сопровождается их созреванием, что включает спирализацию, суперспирализацию (закручивание цепей вокруг гистоновых нуклеосом), химическую модификацию (гидроксилирование, метилирование, гидрирование азотистых оснований) с образованием минорных оснований, соединение с помощью ионных связей с белками, катионами металлов (К+, Са++, Мg++, Мn++, Fе++, Сu++ и т.д.), которые стабилизируют или, наоборот, дестабилизируют вновь синтезированную молекулу.
Синтез и процессинг РНК
В отличии от ДНК, синтез которой происходит в момент деления клетки и служит источником генетической информации для последующих поколений, различные РНК необходимы для генерирования молекул белков. Процесс синтеза рибонуклеиновых кислот с использованием в качестве матрицы фрагментов ДНК называют транскрипцией. Все гены в хромосоме (ДНК с белками) в отсутствии факторов транскрипции находится в выключенном (репрессированном) состоянии, потому что нуклеосомы блокируют области инициирования каждого промотора. Ген будет транскрибироваться после вытеснения нуклеосомы и связывания. Та из двух цепей ДНК, на которой пойдет этот процесс, называется кодирующей. Инициация включает раскручивание участка, разрыв водородных связей, что раскрывает азотистые основания транскриптона, в котором выделяют промотор, собственно ген (единицу транскрипции), терминатор. Первая функциональная единица (промотор) служит для с ним РНК-полимеразы, которая объединяет рибонуклеотиды в последовательность, комплементарную кодирующей цепи гена. Момент терминации распознается терминирующим белком – р-фактором.
Интересно, что в районе промотора расположены сигнальные последовательности двух типов. Одна из них указывает, где должна начаться транскрипция, а другая определяет, как часто должно происходить это событие. Первый локус называют ТАТА-бокс, второй – СААТ-бокс. Кроме них регуляторная зона включает энхансеры или сайленсеры, изменяющие скорость транскрипции, а также гормончувствительные элементы (ГЧЭ), после взаимодействия с которыми кортикостероиды, андрогены, эстрогены, Т3, сАМФ регулируют экспрессию генов.
Практически все первичные транскрипты РНК у эукариот подвергаются сложному процесссингу, т.е. созреванию, начинающемуся в ядре. Будущая иРНК получается после копирования, реакций расщепления, вычленения, легирования, включения дополнительных адениловых нуклеотидов. Ведь свежеполученная РНК содержит неинформативные вставки, считанные с интронов генов, т.е. представляют чередование траслируемых и нетранслируемых участков (гетерогенная ядерная РНК). Процесс удаления интронов и последующего сшивания экзонов получил название сплайсинг (от англ. to splice – соединять концы, сшивать). Он требует наличия специальных ферментов, поэтому осуществляется в крупном рибонуклеопротеидном комплексе – тельце, содержащем РНК и белки, необходимые для эффективного сплайсинга. Эта структура называется сплайсосома. Кроме того к 5'-концу г/яРНК присоединяется 7-метилгуанозинфосфат (кэпирование), который позднее служит стартовой точкой синтеза белка, к 3'-концу надстраивается полиаденилатный хвост (около 200 мононуклеотидов), защищающий иРНК от действия нуклеаз. Параллельно иРНК уже вышедшая из ядра, плотно упаковывается в пространстве.
Первичный транскрипт рРНК не содержит интронов, интенсивно метилируется и расщепляется в ядрышке специфическими РНК-азами на 5S, 18S, 28-рРНК, затем они связываются с белками, образуя рибосомы. Подобным образом (путем частичного гидролиза) осуществляется из первичных транскриптов образование тРНК, формирование вторичной, третичной структуры, химическая модификация. Критическое значение имеет этап сплайсинга в области антикодона, т.к. от него зависит точность выполнения адапторной функции при синтезе белка.
4.4. Синтез полипептидов
Образно говоря, для осуществления двух предыдущих стадий использовался один язык – нуклеотидный, третья же стадия – синтез полипептидов – говорит на другом языке – языке аминокислот. Следовательно, требуется перевод. Поэтому данный процесс и называется трансляцией. Это строительство аминокислотной последовательности на иРНК в качестве матрицы.
Для успешного перевода сведений, записанных в генах ДНК, в строго специфический порядок аминокислот в белке используется биологический (генетический) код.
Положения генетического кода
-
Триплетность. В полипептидах всего 4 разных азотистых основания, аминокислот не менее 20, следовательно – один мононуклеотид не может нести информацию об одной аминокислоте; два (42=16) – тоже, и только количество тринуклеотидов (43=64) может соответствовать числу аминокислот. Причем 3 из них (УАА, УАГ, УГА) бессмысленные (стоп-сигналы) – не несут сведения ни о какой аминокислоте, а 61 – смысловой триплет.
-
Вырожденность. Информация об одной аминокислоте может быть заложена в 1-3, до 5 различных кодонах.
-
Однозначность. С одного кодона можно считать сведения только об одной аминокислоте.
-
Однонаправленность. Считывание производится только в одном направлении с 5'-конца к 3'-концу.
-
Код не перекрывающийся – один и тот же мононуклеотид не может входить в состав двух разных, рядом лежащих кодонов.
-
Код «без запятых» – два рядом расположенных триплета не отделяются друг от друга никакими знаками препинания.
-
Код не универсален, а альтернативен – триплеты ядерной ДНК, несущие сведения об определенных аминокислотах могут отличаться от соответствующих кодонов м-ДНК (см. «Патохимию наследственных болезней», с.18).
Обязательными участниками трансляции служат иРНК, рибосомы, тРНК и аминокислоты. Последние, чтобы стать настоящими субстратами, должны не только активироваться, но и связаться с той тРНК, антикодон которой несет информацию о ней.
Поэтому в клетке присутствуют специальные ферменты, осуществляющие этот процесс и каждый из которых обладает суперспецифичностью, механизм которой называют вторым генетическим кодом. Проверка правильности образования комплекса осуществляется на обеих стадиях, и если выясняется ошибочность, тут же происходит распад образовавшейся структуры:
Е + а/к + АТФ ↔ Е▪аминоацил
АМФ + ФФ
Е▪аминоацил
АМФ + тРНК ↔ Е + АМФ + аминоацилСхематически все процессы образования полипептидов и полинуклеотидов укладываются в следующую схему:
- где над стрелками обозначены названия соответствующих стадий.
Общие принципы реакций
Все биополимеры (полипептиды, гетерополисахариды, полинуклеотиды) имеют организацию, - специфическую упаковку в пространстве, поэтому их синтез включает дополнительно и стадию структурирования. А если их звенья еще и отличаются друг от друга, отсюда, чтобы вновь образующийся полимер включал их в строго определенном порядке, необходима матрица, что и отличает генез белков и нуклеиновых кислот от других подобных процессов. Кроме того, любой синтез требует затрат энергии, причем часто в этих случаях используются не обычные макроэрги, а происходит с их помощью предварительная активация субстратов. Исходя из этих предпосылок, можно выделить следующие общие принципы, характерные для этих процессов:
-
Наличие матрицы: в синтезе дочерней ДНК ею служит вся материнская ДНК; для образования РНК используется фрагмент ДНК (ген, или транскриптон); аминокислотная последовательность определяется набором триплетов иРНК. -
Субстраты предварительно активируются: на первой стадии используются дезоксирибонуклеотидтрифосфаты (дАТФ, дГТФ, дЦТФ, ТТФ), генез РНК требует наличия соответственно АТФ, ГТФ, ЦТФ, УТФ, полипептиды получаются из аминоацил-тРНК. -
Основные ферменты, обеспечивающие собственно полимеризацию, относятся к классу трансфераз и называются ДНК-, РНК-полимеразы и пептидилтрансфераза. -
Сложность образования биополимера выделять в этом процессе две фазы: собственно синтез (как бы начерно) и созревание – укладку в пространстве, химическую модификацию. -
В собственно синтезе выделяют следующие стадии: а) инициацию – начало; б) элонгацию (longus - длинный) – наращивание цепи; в) терминацию – остановку реакций.
Репликация ДНК
Репликация – синтез дочерней ДНК с использованием в качестве матрицы всей молекулы материнской ДНК. С этого процесса и начинается деление клетки в S-фазу (синтетическую). В основе репликации лежат три принципа: а) комплементарности – азотистые основания образующейся дочерней цепи должны обладать химическим и геометрическим соответствием с подобными веществами материнской нити; полуконсервативности – каждая из синтезированных молекул ДНК состоит из одной материнской и одной дочерней; в) однонаправленности – считывание матрицы идет от 5' к 3' концу.
Чтобы порядок азотистых оснований, находящихся внутри нитей ДНК, можно было считывать, необходимо разорвать водородные связи, соединяющие между собой цепи и лишить их спирализации. Для этих целей используются следующие ферменты: топоизомераза и хеликаза. Первая гидролизует на одном из участков полинуклеотида фосфодиэфирная связь, что позволяет этому фрагменту раскрутиться относительно параллельно лежащего:
а затем восстановить связь. Хеликаза разрушает водородные взаимодействия между противолежащими основаниями участков двух цепей:
Под ее действием фрагменты расходятся, образуя репликативную вилку. Затем с помощью праймазы (primer - затравка) синтезируется олигорибонуклеотид – небольшое соединение, состоящее (обратите внимание) из 8-10 монорибонуклеотидов. Оно служит местом, куда крепится основной фермент – ДНК-полимераза. Из-за антипараллельности цепей данные структуры будут локализоваться в разных местах: одна у конца, другая несколько в глубине противоположной нити:
Все дело в том, что ДНК-полимеразы не могут начинать синтез, а способны только добавлять дезоксирибонуклеотидные звенья к 3'-концу уже имеющейся цепи, чем и является праймер. Описано 3 класса данных энзимов: в ядрах присутствуют полимеразы альфа (Роl α), ответственные за хромосомную репликацию, полимеразы бета (Роl β), которые используются при необходимости при репарации. Третий их представитель – ДНК-полимераза гамма (Роl γ) осуществляет синтез кольцевого генома митохондрий.
Присоединение каждого нового нуклеотидного остатка к 3'-концу растущей цепи сопровождается гидролизом макроэргической связи в дНТФ и отщеплением пирофосфата.
Важная деталь – генетический материал живых организмов имеет огромные размеры, но реплицируется с высокой точностью. В среднем при воспроизведении генома (ДНК длиной более 3 млрд пар нуклеотидов) возникает не более 3-х ошибок благодаря наличию специальных механизмов, осуществляющих необходимую коррекцию. Суть последней в том, что ДНК-полимеразы дважды проверяют соответствие каждого нуклеотида матрице: один раз перед включением его в состав растущей цепи и второй – перед присоединением нового нуклеотида. Очередная фосфодиэфирная связь синтезируется лишь в том случае, если последний мономер удлиняющейся дочерней нити ДНК образовал правильную (комплементарную) пару с соответствующим нуклеотидом матрицы. Если же на предыдущей стадии произошло ошибочное спаривание оснований, то фермент возвращает последнее добавленное звено, после чего освободившееся место занимает правильный нуклеотид.
Параллельно с ростом цепи продолжается деспирализация и разделение цепей с помощью топоизомеразы и хеликазы, удлинение репликативной вилки. А так как процесс осуществляется лишь в одном (5' – 3') направлении, что позволяет происходить этому явлению непрерывно только на одной из матричных нитей. На антипараллельной синтез ограничивается сравнительно короткими фрагментами (100-1000 мононуклеотидов), названными по имени обнаружившего их ученого фрагментами Оказаки (с постоянным предварительным синтезом праймеров):
Так продолжается до тех пор, пока вся матрица не выполнит свою функцию, причем во время репликации специальные нуклеазы вычленяют праймеры. Если позволяет направление, ДНК-полимеразы застраивают возникшие бреши, а сшивание сближенных фрагментов обеспечивается ДНК-лигазами – так завершается собственно синтез (стадия терминации).
В тех концах дочерних цепей, где находились праймеры, которые позднее были удалены, не происходит достраивания дезоксирибонуклеотидами (невозможно считывание в обратном направлении), отсюда при каждом делении клетки молекулы новых цепей укорачиваются на 10-20 нуклеотидов, но объем информации при этом не уменьшается, так как потерянные участки ее не несли. Дело в том, что когда в эмбриональных тканях шли усиленные митозы параллельно с этим осуществлялись обратные транскрипции. С помощью теломеразы концы ДНК наращивались теломерными повторами ТТАGGG, роль матрицы при этом выполняла РНК, включающая 450 мононуклеотидов. Поскольку многократно повторенные теломерные последовательности не являются кодирующими, их утрата в процессе деления не приводит к потере информативных участков, тем самым обеспечивается передача генетического материала в поколениях клеток.
Однако в родившемся организме активность теломераз регистрируется лишь в половых, раковых, стволовых клетках, в соматических же эти ферменты не работают. Теломеры постепенно укорачиваются, что в конце концов запускает процессы остановки клеточного цикла и провоцирует апоптоз. Интересный факт: ученые, исследующие рак, до сих пор используют культуру клеток Неlа из опухоли женщины по имени Генриетта Лакс. Эта больная умерла в 1951 году, но клетки ее новообразования продолжают делиться и собираются жить вечно.
Происходящее в момент деления клетки удлинение синтезирующихся нитей ДНК сопровождается их созреванием, что включает спирализацию, суперспирализацию (закручивание цепей вокруг гистоновых нуклеосом), химическую модификацию (гидроксилирование, метилирование, гидрирование азотистых оснований) с образованием минорных оснований, соединение с помощью ионных связей с белками, катионами металлов (К+, Са++, Мg++, Мn++, Fе++, Сu++ и т.д.), которые стабилизируют или, наоборот, дестабилизируют вновь синтезированную молекулу.
Синтез и процессинг РНК
В отличии от ДНК, синтез которой происходит в момент деления клетки и служит источником генетической информации для последующих поколений, различные РНК необходимы для генерирования молекул белков. Процесс синтеза рибонуклеиновых кислот с использованием в качестве матрицы фрагментов ДНК называют транскрипцией. Все гены в хромосоме (ДНК с белками) в отсутствии факторов транскрипции находится в выключенном (репрессированном) состоянии, потому что нуклеосомы блокируют области инициирования каждого промотора. Ген будет транскрибироваться после вытеснения нуклеосомы и связывания. Та из двух цепей ДНК, на которой пойдет этот процесс, называется кодирующей. Инициация включает раскручивание участка, разрыв водородных связей, что раскрывает азотистые основания транскриптона, в котором выделяют промотор, собственно ген (единицу транскрипции), терминатор. Первая функциональная единица (промотор) служит для с ним РНК-полимеразы, которая объединяет рибонуклеотиды в последовательность, комплементарную кодирующей цепи гена. Момент терминации распознается терминирующим белком – р-фактором.
Интересно, что в районе промотора расположены сигнальные последовательности двух типов. Одна из них указывает, где должна начаться транскрипция, а другая определяет, как часто должно происходить это событие. Первый локус называют ТАТА-бокс, второй – СААТ-бокс. Кроме них регуляторная зона включает энхансеры или сайленсеры, изменяющие скорость транскрипции, а также гормончувствительные элементы (ГЧЭ), после взаимодействия с которыми кортикостероиды, андрогены, эстрогены, Т3, сАМФ регулируют экспрессию генов.
Практически все первичные транскрипты РНК у эукариот подвергаются сложному процесссингу, т.е. созреванию, начинающемуся в ядре. Будущая иРНК получается после копирования, реакций расщепления, вычленения, легирования, включения дополнительных адениловых нуклеотидов. Ведь свежеполученная РНК содержит неинформативные вставки, считанные с интронов генов, т.е. представляют чередование траслируемых и нетранслируемых участков (гетерогенная ядерная РНК). Процесс удаления интронов и последующего сшивания экзонов получил название сплайсинг (от англ. to splice – соединять концы, сшивать). Он требует наличия специальных ферментов, поэтому осуществляется в крупном рибонуклеопротеидном комплексе – тельце, содержащем РНК и белки, необходимые для эффективного сплайсинга. Эта структура называется сплайсосома. Кроме того к 5'-концу г/яРНК присоединяется 7-метилгуанозинфосфат (кэпирование), который позднее служит стартовой точкой синтеза белка, к 3'-концу надстраивается полиаденилатный хвост (около 200 мононуклеотидов), защищающий иРНК от действия нуклеаз. Параллельно иРНК уже вышедшая из ядра, плотно упаковывается в пространстве.
Первичный транскрипт рРНК не содержит интронов, интенсивно метилируется и расщепляется в ядрышке специфическими РНК-азами на 5S, 18S, 28-рРНК, затем они связываются с белками, образуя рибосомы. Подобным образом (путем частичного гидролиза) осуществляется из первичных транскриптов образование тРНК, формирование вторичной, третичной структуры, химическая модификация. Критическое значение имеет этап сплайсинга в области антикодона, т.к. от него зависит точность выполнения адапторной функции при синтезе белка.
4.4. Синтез полипептидов
Образно говоря, для осуществления двух предыдущих стадий использовался один язык – нуклеотидный, третья же стадия – синтез полипептидов – говорит на другом языке – языке аминокислот. Следовательно, требуется перевод. Поэтому данный процесс и называется трансляцией. Это строительство аминокислотной последовательности на иРНК в качестве матрицы.
Для успешного перевода сведений, записанных в генах ДНК, в строго специфический порядок аминокислот в белке используется биологический (генетический) код.
Положения генетического кода
-
Триплетность. В полипептидах всего 4 разных азотистых основания, аминокислот не менее 20, следовательно – один мононуклеотид не может нести информацию об одной аминокислоте; два (42=16) – тоже, и только количество тринуклеотидов (43=64) может соответствовать числу аминокислот. Причем 3 из них (УАА, УАГ, УГА) бессмысленные (стоп-сигналы) – не несут сведения ни о какой аминокислоте, а 61 – смысловой триплет. -
Вырожденность. Информация об одной аминокислоте может быть заложена в 1-3, до 5 различных кодонах. -
Однозначность. С одного кодона можно считать сведения только об одной аминокислоте. -
Однонаправленность. Считывание производится только в одном направлении с 5'-конца к 3'-концу. -
Код не перекрывающийся – один и тот же мононуклеотид не может входить в состав двух разных, рядом лежащих кодонов. -
Код «без запятых» – два рядом расположенных триплета не отделяются друг от друга никакими знаками препинания. -
Код не универсален, а альтернативен – триплеты ядерной ДНК, несущие сведения об определенных аминокислотах могут отличаться от соответствующих кодонов м-ДНК (см. «Патохимию наследственных болезней», с.18).
Обязательными участниками трансляции служат иРНК, рибосомы, тРНК и аминокислоты. Последние, чтобы стать настоящими субстратами, должны не только активироваться, но и связаться с той тРНК, антикодон которой несет информацию о ней.
Поэтому в клетке присутствуют специальные ферменты, осуществляющие этот процесс и каждый из которых обладает суперспецифичностью, механизм которой называют вторым генетическим кодом. Проверка правильности образования комплекса осуществляется на обеих стадиях, и если выясняется ошибочность, тут же происходит распад образовавшейся структуры:
Е + а/к + АТФ ↔ Е▪аминоацил