Файл: ан айналымы биохимиясы модулі бойынша апаратты топтама.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 12.12.2023
Просмотров: 357
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
СОДЕРЖАНИЕ
Жоғарыда айтылған тотығу стрессі көптеген жүрек-қан тамырлары патологиясының белгісі ретінде қызмет етеді. Реактивті оттегі түрлерінің көбеюі қант диабетіне, атеросклерозға және көптеген артериялық гипертензияға тән. Бұл жағдайларда ренин-ангиотензин жүйесінің жоғары белсенділігі жиі байқалады, ал ангиотензин II тотығу стрессінің күшті қоздырғышы болып табылады, ол бір жағынан eNOS белсенділігін төмендетеді (мысалы, тотыққан төмен тығыздықты липопротеидтер C протеині С-ны белсендіре алады), ал екінші жағынан гендердің экспрессиясын төмендетеді. eNOS, бұл сонымен қатар NO өндірісін азайтады.
Антиоксиданттарды немесе ангиотензин II (ангиотензинді түрлендіретін фермент ингибиторлары немесе ангиотензин II блокаторлары) түзілуіне немесе әсеріне кедергі келтіретін заттарды қолдану NO түзілуін әрдайым арттырады. Ауру кезінде азот оксиді өндірісінің төмендеуі тек eNOS-қа тікелей әсер етумен байланысты емес деп айту керек. Осылайша, глюкокортикоидтардың эндотелийге әсері ферменттің өзін ғана емес, оның кофакторы - тетрагидробиоптеринді де төмендетеді.
Эндотелий NO синтазасының жұмысын бұзу оның негізгі субстратының, L-аргининнің жетіспеуімен байланысты болуы мүмкін. Әдетте, бұл аминқышқыл организмге тамақпен бірге жеткілікті мөлшерде түседі және сонымен қатар ересек организмде тікелей синтезделуі мүмкін. Алайда, NO синтазаларынан басқа, аргинин көптеген басқа ферменттер үшін субстрат қызметін атқарады, атап айтқанда, тамырлардың эндотелийін қоса, әр түрлі жасушаларда орналасқан аргиназа. Қант диабетінде, тотығу стрессінде, сондай-ақ иммундық жүйенің жасушалары бөлетін цитокиндердің әсерінен қабыну процестерінде (ісік некрозы факторы және т.б.) эндотелийдегі аргиназа мөлшері артады.
Соңында, эндотелиальды NO синтаза тежегіштері, мысалы, диметиларгинин, адамдар мен басқа жануарларда пайда болуы мүмкін. Эндотелий NO синтазасының бұл «жалған субстраты» ферменттің белсенді алаңы үшін L-аргининмен шынайы субстратпен бәсекелеседі. Әдетте диметиларгинин организмде аз мөлшерде ғана қалыптасады (ересек адамда, тәулігіне 60 мг), алайда қан айналымы патологиясының алуан түрлілігімен (артериялық гипертензия, атеросклероз, коронарлық жеткіліксіздік және т.б.) оның өндірісі айтарлықтай артады, ал эндотелий NO белсенділігі -синтаза сәйкесінше азаяды. Демек, азот оксиді эндотелийдің іргелес тегіс бұлшықет жасушаларына босаңсыту әсерін тигізетін маңызды реттеуші фактор болып табылады
, вазодиляцияны тудырады және жүйелік деңгейде қан қысымының қажетсіз жоғарылауын тегістейді. Эндотелий осы мәселелерді шешуге жеткілікті мөлшерде NO бөлу қабілетін сақтаған кезде, қан тамырлары жүйесінің күйі туралы алаңдаудың қажеті жоқ.
РААЖ, кининдер, простаноидтар
Біздің денеде протеолитикалық ферменттердің өзара байланысқан екі жүйесі бар, олардың жұмысының нәтижесінде тамырлы тонус реттеледі.
1. РЕНИН-АНГИОТЕНЗИН-АЛЬДОСТЕРОДЫ ЖҮЙЕ (РААЖ-жүйесі).
Бұл жүйенің жұмысы қан қысымын арттыруға бағытталған.
2. КИНИН ЖҮЙЕСІ. Қан қысымын төмендетуге бағытталған.
Екі жүйенің де активтенуі шектеулі протеолиз реакцияларымен олардың прекурсорларынан биологиялық белсенді төмен молекулалы пептидтердің синтезіне дейін азаяды.
Негізгі рөл тамырлар тонусын және су-тұз алмасуын реттейтін РААЖ-қа тиесілі.
Бүйректе протеолитикалық фермент - РЕНИН джекстагломерулярлық аппараттың жасушаларында (ЮГА) синтезделеді. Ренин ангиотензиногенді декапептидті ангиотензин-I-ге шектеулі протеолизге айналдырып, тамырлар тонусын реттеуге қатысады. Ангиотензин-I-ден, карбоксикатепсин ферментінің әсерінен октапептид ангиотензин-II түзіледі (сонымен қатар шектеулі протеолиз арқылы).
Ол вазоконстрикторлы әсер етеді, сонымен қатар бүйрек үсті безінің кортекс гормоны - альдостерон өндірісін ынталандырады. Альдостерон бүйрек түтікшелеріндегі натрий мен судың реабсорбциясын күшейтеді - бұл тамырларда айналымдағы қан көлемінің ұлғаюына әкеледі. Нәтижесінде қан қысымы көтеріледі. Ангиотензин II молекуласы өз қызметін атқарған кезде арнайы протеиназалар тобы - ангиотензиназалар арқылы толық протеолизге ұшырайды. РЕНИН-АНГИОТЕНСИН-АЛДОСТЕРОН ЖҮЙЕСІ осылай жұмыс істейді.
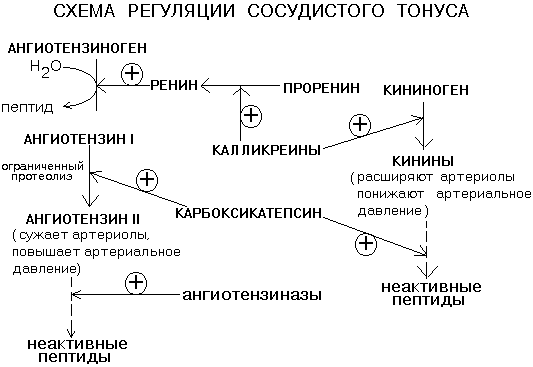
Ренин өндірісі бүйректің қанмен қамтамасыз етілуіне байланысты. Сондықтан қан қысымының төмендеуімен ренин өндірісі жоғарылайды, ал жоғарылаған сайын ол азаяды. Бүйрек патологиясы кезінде рениннің жоғарылауы кейде байқалады және тұрақты гипертензия (артериялық қысымның жоғарылауы) дамуы мүмкін. Ренин-ангиотензин-альдостерон жүйесі тамырлы тонусты реттейтін басқа жүйемен тығыз байланыста жұмыс істейді: КАЛЛИКРЕЙН-КИНИН ЖҮЙЕСІ, оның әрекеті қан қысымының төмендеуіне әкеледі.
Кининоген ақуызы бүйректе синтезделеді. Қанға түскеннен кейін кининоген серин протеиназы - калликреиндер арқылы вазоактивті пептидтерге - кининдерге айналады: брадикинин және каллидин. Брадикинин мен каллидиннің вазодилатациялық әсері бар - олар қан қысымын төмендетеді. Кининдерді инактивациялау карбоксикатепсиннің қатысуымен жүреді - бұл фермент бір мезгілде тамырлық тонустың екі реттелу жүйесіне де әсер етеді, бұл атериялық қысымның жоғарылауына әкеледі. Карбоксикатепсин ингибиторлары гипертонияның кейбір түрлерін емдеуде дәрілік жолмен қолданылады.
Кинин жүйесінің ақуыздары кининдердің 6 түрі анықталды:
1. Нонапептидті брадикинин
2. Каллидин
3. Мел-Лис-брадикинин
4. Т-кинин
5. Р заттар
6. Энпирокининдер (ВМК және НМК, кининогендер түрінде шығарылады)
Кининдер кинин гормондары немесе жергілікті гормондар деп аталады. Олар ішкі секреция бездерінде түзілмейді, бірақ белсенді емес қосылыстардан - кининогендерден бөлінеді, олар негізінен бауырда синтезделеді, өкпеде, бүйректе, сондай-ақ теріде және дәнекер тіндерінде де синтезделуі мүмкін. Кининогендердің кининдерге айналуы арнайы ферменттердің – кининогеназалардың (калликреиндердің), әсерінен жүреді. Калликреиндер бауырда калликреиноген түрінде синтезделеді, ол өз кезегінде Хагеман факторы, адреналин, трипсин, урокиназа немесе қан қышқылдануы кезінде белсендіріледі және калликреин-белсенді ферментіне айналады.
Калликреин кининогендерді белсендіреді: брадикининоген брадикининге және каллидиноген каллидинге айналады. Олардың әрекеті қан тамырларының тегіс бұлшықеттеріне және капиллярлар мембранасына бағытталған. Кинин жүйесінің нәруыздары қан плазмасындағы кининаза әсерінен инактивтенеді. Қандағы кининдердің мөлшері өте аз және литріне наномольмен өлшенеді.
Кининдердің рөлі:
A) Олар қан тамырлар қабырғасының тонусына тікелей әсер етіп, прекапиллярлардың кеңеюіне және мембраналардың өткізгіштігінің жоғарылауын тудырады, осыған байланысты олар гемодинамиканың реттелуіне қатысады, өйткені тегіс бұлшықеттерге әсер етеді. Кининдер капиллярлар мен тәждік тамырларды кеңейтіп, гипотензиялық әсер береді. Гипертония патогенезінде кинин синтезінің бұзылуы болуы ықтимал. Кининдер аз мөлшерде әрекет етеді. Қан қысымының төмендеуін тудыратын брадикининнің минималды дозасы 0,1 мкг / кг құрайды. Сонымен қатар, брадикинин капиллярларға әсер ете отырып, олардың мембраналарының өткізгіштігін күрт арттырады, бұл ісінуге, гемостазға және басқа қабыну белгілеріне ықпал етуі мүмкін. Капиллярлық мембраналар брадикининнің шамадан тыс мөлшерінің әсерінен «елекке» айналады, оның саңылауы арқылы оның компоненттері қан ағымынан шығады. Салицилаттар (мысалы, аспирин) брадикинин түзілуін тежейтіні белгілі, бұл қабынудың төмендеуімен қатар жүреді. Бұл осы препараттарды қабыну ауруларын емдеуде қолдануға негіз болады.
B) Брадикинин атеросклероздық түйіндердің пайда болуына әсер етеді, олар көбінесе тамырлар қабырғасында «саңылаулар» бар жерлерде жиналады. «Саңылаулардың» пайда болуы қан тамыр қабырғасының кеңеюімен байланысты
Брадикинин:
гистаминді мастоциттерден босатады;
әртүрлі тіндердегі цитокиндердің синтезін ынталандырады және оларды босатып шығарады;
бірқатар интерлейкиндерді босатып шығарады;
репарация процестеріне қатысады;
перифериялық тіндердің глюкозаны қабылдауын ынталандырып, инсулинге ұқсас әсер етеді;
орталық және перифериялық жүйке жүйесінде жүйке импульстарының берілуін модуляциялайды, гематоэнцефалдық тосқауылдың күйін өзгертеді;
қан қысымын төмендетеді;
капиллярлардың өткізгіштігін жоғарылатады.
Бронхтардың және басқа органдардың тегіс бұлшықеттерін азайтады. Кининдер бронхтың жиырылуына ықпал ететін және бронх демікпесін тудыратын бронхоконстрикторлық әсерге ие. Сондықтан осы патологияны емдеу үшін кининазаның активтенуін тудыратын дәрілерді, яғни кининдердің ыдырауына ықпал ететін ферменттер іздеу жүргізіледі.
Лейкоциттердің диапедезін ынталандырады және ауырсыну эффектісін тудырады.
Брадикинин, кем дегенде, екі түрлі арнайы рецепторлармен - В1 рецепторымен және В2 рецепторымен әрекеттескенде көптүрлі биологиялық әсер көрсетеді.
D) Кининдер жүректің соғу жиілігін және күшін жоғарылатады, О2 пайдалануын көбейтеді және осы тіндегі метаболизмді күшейтеді
E) Кининдер ойық жара ауруының (HCl секрециясын күшейтеді) патогенезінде маңызды рөл атқарады.
Простаноидтардың қан тамырлар тонусына әсері
Простагландиндер (PGs) - шығу тегі бойынша арахидон қышқылымен байланысты физиологиялық белсенді заттар, және бұл заттар тобын эйкозаноидтар деп атайды. ПГ тіндердің метаболизмі процесінде бөлінеді және басқа эйкозаноидтардан өзгешелігі, олар қан тамыр тонусына әсер ете алады. Дәл осындай вазоактивті мөлшері қан тамырларын қоршаған жасушадан тыс ортада олардың қалыпты жұмыс істеуі жағдайында түзілетінін атап өту маңызды. Қарастырылған топтағы заттардың қан тамырларына әсер ету қашықтығы үлкен емес. Е типті ПГ-дер өте төмен концентрацияда барлық органдарда қан ағымына тамырлық төзімділіктің төмендеуін тудыруы мүмкін. Қарқынды жұмыс жасайтын бұлшықеттердің веноздық қанындағы PG мөлшері жоғарылайды және бұл осы фактордың жұмыс гиперемиясының дамуына қатысуын болжайды.
ПГ трансмиттердің (норадреналиннің) жүйке ұштарынан шығуын реттеудегі рөлі дәлелденген. Сонымен, Е тобы ПГ норадреналиннің бөліну мөлшерін де, оның рецепторларға әсерін де тежейді, ал F тобы ПГ керісінше әсер етеді. Демек, ПГ тамыр тонусын жүйкелік бақылауға қатысады. А және Е класындағы простагландиндер, сондай-ақ простациклин қан тамырларын кеңейтіп, ағзалар мен тіндерде қан айналымын жақсартады. F класындағы простагландиндер мен тромбоксандар қан тамырларын тарылтып, тромбоциттер агрегациясын тудырады.
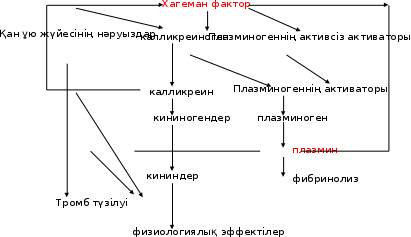
Коагуляция жүйесінің болуына қарамастан, организмдегі қан сұйық күйде болады, өйткені коагуляция факторларынан басқа, оның коагуляциясын болдырмайтын заттар - қан ұюға қарсы жүйе бар. Физиологиялық жағдайда бұл жүйелер теңестірілген. Қанның құрамында қан ұйындысының (тромбтың) еруін (фибринолизді) қамтамасыз ететін нәруыздар бар. Бұл процесс плазминогенді белсендіру арқылы жүзеге асырылады, ол плазминге айналады, соңғысы фибриногеннің де, фибриннің де ыдырауын тудырады. Кининдердің түзілуі, қан ұюы және фибринолиз арасында тығыз байланыс бар, себебі олардың ортақ активаторлары - Хагеман факторы және плазмин бар.
Жоғарыда айтылғандай, Хагеман факторы қанның коагуляциясының бастаушы механизмі болып табылады, сонымен бірге ол калликреиногеннің калликреинге айналуын белсендіреді және сондықтан кининдердің түзілуіне ықпал етеді, сонымен қатар белсенді емес плазминогеннің белсенді плазминге айналуын тудырады. Плазмин калликреиннің түзілуіне қатысады, белсенді ферменттің - XII фактордың түзілуіне ықпал етеді. Калликреиноген - плазминоген мен XII фактордың активаторы. Плазмин тромбиннің түзілуін тежейді, ал калликреин фибриноген активациясының ингибиторы болып табылады.
ЛИМФА
Қан тамырларына күніне 1-ден 2 литрге дейін лимфа түседі. Лимфа құрамы лимфа тамырларының анатомиялық құрылымы мен орналасуына байланысты және әр адам үшін жеке болып табылады. Лимфада қанмен салыстырғанда эритроциттер өте аз, ал лимфоциттер көп. Лимфадағы нәруыз мөлшері 0,2-ден 6% -ке дейін болуы мүмкін. Альбуминдер мен глобулиндердің арақатынасы = 4/1. Бұл қан плазмасына қарағанда әлдеқайда жоғары.
Осмостық қысымды сақтаудағы натрий, хлор және
басқа электролиттердің рөлі