Файл: Воронова, Климова, Менджерицкий Анатомия центральной нервной системы.pdf
Добавлен: 19.02.2019
Просмотров: 4152
Скачиваний: 26

Рис. 50. Пирамидный нейрон коры:
1 — апикальный дендрит; 2 — тело нейрона; 3 — горизонтальный дендрит; 4 — аксон
♦ III слой — пирамидный (lamina pyramidalis),
♦ III слой — пирамидный (lamina pyramidalis), толщиной около 1 мм, состоит из мелких и средних
пирамидных клеток. Типичный пирамидный нейрон имеет форму треугольника, вершина которого
направлена вверх (рис. 50). От вершины отходит апикальный дендрит, ветвящийся в
вышележащих слоях. Аксон пирамидной клетки отходит от основания клетки и направляется
вниз. Дендриты клеток III слоя направляются во второй слой. Аксоны клеток III слоя
оканчиваются на клетках нижележащих слоев или образуют ассоциативные волокна.
♦ IV слой — внутренний зернистый (lamina granulans internus).
♦ IV слой — внутренний зернистый (lamina granulans internus). Он состоит из звездчатых клеток,
имеющих короткие отростки, и малых пирамид. Дендриты клеток IV слоя уходят в молекулярный
слой коры, а их коллатерали ветвятся в своем слое. Аксоны клеток IV слоя могут подниматься в
вышележащие слои или уходить в белое вещество как ассоциативные волокна. Толщина IV слоя
от 0,12 до 0,3 мм.
♦ V слой — ганглионарный (lamina ganglionaris)
♦ V слой — ганглионарный (lamina ganglionaris) — слой больших пирамид. Самые крупные клетки
коры расположены именно в этом слое (гигантские пирамиды Беца передней центральной
извилины) (см. рис. 49Б). Их апикальные дендриты достигают молекулярного слоя, а базальные
дендриты распределяются в своем слое. Аксоны клеток V слоя покидают кору и являются
ассоциативными, комиссуральными или проекционными волокнами. Толщина V слоя достигает
0,5 мм.
93
♦ VI слой коры — полиморфный (lamina multiformis).
♦ VI слой коры — полиморфный (lamina multiformis). Содержит клетки разнообразной формы и
размера, имеет толщину от 0,1 до 0,9 мм. Часть дендритов клеток этого слоя достигает
молекулярного слоя, другие же остаются в пределах IV и V слоев. Аксоны клеток VI слоя могут
подниматься к верхним слоям или уходить из коры в качестве коротких или длинных
ассоциативных волокон (см. рис. 49).
Клетки одного слоя коры выполняют сходную функцию в обработке информации. I и IV слои
являются местом ветвления ассоциативных и комиссуральных волокон, т.е. получают
информацию от других корковых структур. III и IV слои являются входными, афферентными для
проекционных полей, так как именно в этих слоях заканчиваются таламические волокна. V слой
клеток выполняет эфферентную функцию, его аксоны несут информацию к нижележащим
структурам мозга. VI слой также является выходным, но его аксоны кору не покидают, а являются
ассоциативными.
Основным принципом функциональной организации коры является объединение нейронов в
колонки. Колонка расположена перпендикулярно поверхности коры и охватывает все ее слои от
поверхности к белому веществу. Связи между клетками одной колонки осуществляются по
вертикали вдоль оси колонки. Боковые отростки клеток имеют небольшую длину. Связь между
колонками соседних зон осуществляется через волокна, уходящие вглубь, а затем входящие в

другую зону, т.е. короткие ассоциативные волокна. Функциональная организация коры в виде
колонок обнаружена в соматосенсорной, зрительной, моторной и ассоциативной коре.
Отдельные зоны коры имеют принципиально одинаковое клеточное строение, однако есть и
отличия, особенно в структуре III, IV и V слоев, которые могут распадаться на несколько
подслоев. Кроме этого, существенными цитоархитектоническими признаками являются плотность
расположения и размеры клеток, наличие специфических типов нейронов, расположение и
направление хода миелиновых волокон (см. рис. 49В).
Цитоархитектонические особенности позволили разделить всю поверхность коры на 11
цитоархитектонических областей, включающих в себя 52 поля (по Бродману). Каждое
цитоархитектоническое поле обозначено на картах мозга номером, который присваивался ему в
порядке описания. Следует отметить, что между цитоархитектоническими полями не существует
резких границ,
94
Рис. 51. Латеральная поверхность левого полушария (Сильвиева борозда
раздвинута):
1 — центральная борозда; 2 — теменная доля; 3 — задняя центральная извилина (соматосенсорная кора); 4
— ассоциативные зрительные зоны; 5 — латеральная (Сильвиева) борозда; 6 — первичное зрительное поле
(участок зрительного восприятия); 7 — вторичное зрительное поле; 8 — первичный слуховой центр; 9 —
вторичный слуховой центр с речевым центром Вернике; 10 — ассоциативные слуховые зоны; 11 — височная
доля; 12 — островок (находится в глубине Сильвиевой борозды); 13 — двигательный речевой центр (центр
Брока) — при праворукости находится только в левом полушарии; 14 — лобная доля мозга; 15 —
прецентральная извилина (двигательные зоны)
клеточные слои плавно меняют свою структуру при переходе от одного поля к другому (рис. 51).
Каждое поле коры выполняет определенную функцию. Часть полей коры являются сенсорными. В
первичных сенсорных полях заканчиваются проекционные афферентные волокна. Из первичных
сенсорных полей информация по коротким ассоциативным волокнам передается во вторичные
проекционные поля, расположенные рядом с ними. Так, поля 1 и 3, занимающие медиальную и
латеральную поверхность задней центральной извилины, являются первичными проекционными
полями кожной чувствительности противоположной половины поверхности тела. Расположенные
рядом друг с другом участки кожи также рядом друг с другом проецируются на корковой
поверхности. Такая организация проекций называется топической. В медиальной части
представлены нижние конечности, а наиболее низко на латеральной части извилины расположены
проекции рецепторных полей кожной поверхности головы. При этом участки поверхности тела,
богато снабженные рецепторами (пальцы, губы, язык), проецируются на боль-
95
шую площадь коры, чем участки, имеющие меньшее количество рецепторов (бедро, спина, плечо).
Поле 2, расположенное в нижнелатеральной части той же извилины, является вторичным
проекционным полем кожной чувствительности.
Поля 17—19, расположенные в затылочной доле, являются зрительным центром коры, 17-е поле,
занимающее сам затылочный полюс, является первичным. Прилежащие к нему 18-е и 19-е поля
выполняют функцию вторичных ассоциативных полей и получают входы от 17-го поля.
В височных долях расположены слуховые проекционные поля (41, 42). Рядом с ними на границе
височной, затылочной и теменной долей расположены 37-е, 39-е и 40-е, характерные только для
коры головного мозга человека. У большей части людей в этих полях левого полушария
расположен центр речи, отвечающий за восприятие устной и письменной речи. Поле 43,
занимающее нижнюю часть задней центральной извилины, получает вкусовые афференты.
Структуры обонятельной чувствительности посылают свои сигналы в кору больших полушарий
без переключений в других отделах ЦНС. Обонятельные луковицы расположены под нижней
поверхностью лобных долей (см. рис. 55, 7). От них начинается обонятельный тракт, который
является первой парой черепно-мозговых нервов (п. olfactorius). Корковыми проекциями
обонятельной чувствительности являются структуры древней коры (рис. 52).
Двигательные области коры расположены в прецентральной извилине лобной доли (впереди от
проекционных зон кожной чувствительности) (рис. 52, 1). Эту часть коры занимают поля 4 и 5. Из
V слоя этих полей берет начало пирамидный путь, заканчивающийся на мотонейронах спинного
мозга. Расположение и соотношение зон иннервации сходно с проекционным представительством
кожного анализатора, т.е. имеет соматотопическую организацию. В медиальных частях извилины
расположены колонки, регулирующие деятельность мышц ног, в нижней части, у латеральной
борозды — мышц лица и головы противоположной стороны тела.
Афферентные и эфферентные проекционные зоны коры занимают относительно небольшую ее
площадь. Большая часть поверхности коры занята третичными или межанализаторными зонами,
называемыми ассоциативными.
Ассоциативные зоны коры занимают значительное пространство между лобной, затылочной и
височной корой (60-70% новой коры). Они получают полимодальные входы от сенсорных облас-
96
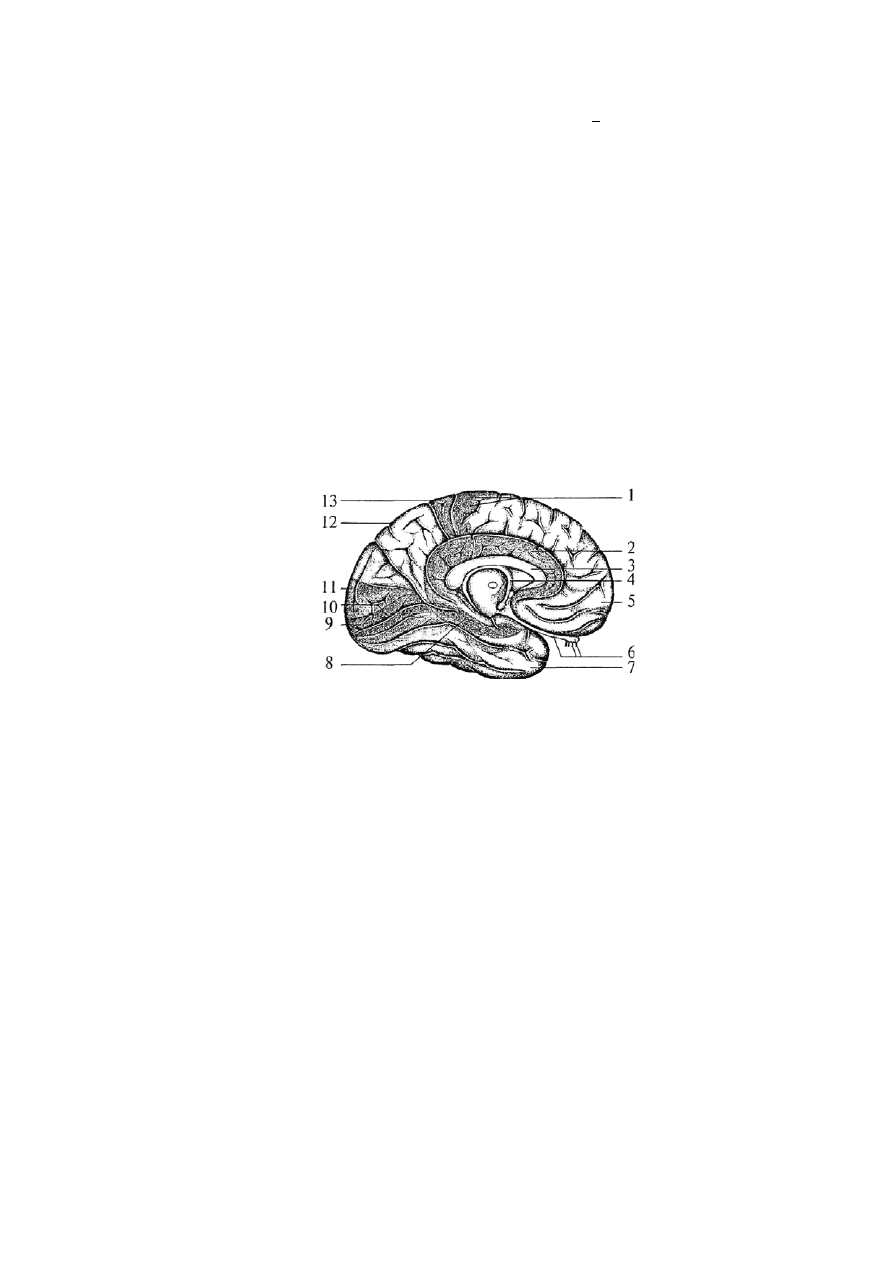
Рис. 52. Медиальная поверхность левого полушария:
1 — прецентральная извилина (двигательные зоны); 2 — поясная извилина (часть лимбической системы),
отвечает за висцеральную чувствительность; 3 — мозолистое тело (основная комиссура); 4 — свод; 5 —
лобная доля; 6 — обонятельные нервы, обонятельная луковица и обонятельный путь; 7 — височная доля; 8
— гиппокамп (часть лимбической системы); 9 — первичное проекционное зрительное поле (17 поле); 10 —
вторичное проекционное зрительное поле (18 поле);
11 — затылочная доля; 12 — теменная доля; 13 — задняя центральная извилина (соматосенсорные зоны)
тей коры и таламических ассоциативных ядер и имеют выходы на двигательные зоны коры.
Ассоциативные зоны обеспечивают интеграцию сенсорных входов и играют существенную роль в
процессах высшей нервной и психической деятельности.
Филогенез.
Филогенез. У низших позвоночных (круглоротые и рыбы) развитие конечного мозга идет по пути
утолщения основания, в котором формируются базальные ганглии в виде больших парных
выпуклых масс. Боковые стенки и крыша конечного мозга, образующие плащ (мантию), остаются
тонкими и повторяют форму полосатых тел в виде двух полушарий, хотя на два полушария
конечный мозг еще не разделен, не разделена еще и полость его желудочка. В передней стенке
конечного мозга развивается парное выпячивание — обонятельные доли. Они продолжаются в
более или менее длинный стебелек, заканчивающийся обонятельной луковицей. Серое вещество
мантии выстилает полость желудочка, а ее поверхность содержит белое вещество. В концевой
пластинке располагается передняя комиссура, связывающая между собой полосатые тела.
У двоякодышащих рыб и первых наземных позвоночных (амфибии) передняя стенка мозгового
пузыря вдвигается посередине между полосатыми телами в виде продольной складки, разделяю-
97
щей конечный мозг на два полушария. Полость мозгового пузыря разделяется на два боковых
желудочка, которые соединяются сзади Монроевыми отверстиями с III желудочком.
Мантия еще тонкая, но нервные клетки перемещаются к поверхности, однако остаются под слоем
белого вещества. Над передней комиссурой появляется еще одна комиссура, соединяющая между
собой мантийные структуры полушарий (аналог комиссуры гиппокампа).
У рептилий конечный мозг разрастается еще больше, но это разрастание идет в основном за счет
развития базальных ганглиев. Мантия остается тонкой, но на ее поверхности впервые появляется
серое вещество, которое представляет собой кору. Эта кора является высшей обонятельной
структурой и сравнима с гиппокампом млекопитающих.
Усложнение организации конечного мозга млекопитающих происходит за счет развития мантии.

Мантия покрыта корой. Кроме древней и старой коры в филогенетическом ряду млекопитающих
появляется новая кора. Чем более высокоразвит вид млекопитающих, тем большую поверхность
занимают структуры новой коры. Вместе с развитием мантии преобразуются и полости
желудочков.
В добавление к имеющимся комиссурам появляется мозолистое тело, связывающее структуры
новой коры обоих полушарий. С увеличением поверхности новой коры нарастает количество
волокон мозолистого тела и его мощность. Мозолистое тело значительно разрастается и сдвигает
назад комиссуру гиппокампа.
Кора у низших млекопитающих развивается в связи с обонятельной сенсорной системой. У
высших млекопитающих (обезьяны, человека) с обонянием связаны только структуры древней и
старой коры.
Кроме того, уже на ранних этапах развития позвоночных (амфибии и рептилии) кора получает
афферентацию от сенсорных систем других модальностей через ядра таламуса. У млекопитающих
кроме специфических ядер таламуса возникают и развиваются ассоциативные ядра, имеющие
связи с неокортексом и определяющие развитие конечного мозга.
Третьим источником афферентных воздействий на кору больших полушарий является
гипоталамус, который играет роль высшего регуляторного центра вегетативных функций. У
млекопитающих филогенетически более древние отделы переднего гипоталамуса связаны с
древней формацией коры и с древними ядрами амигдалы. Молодые отделы заднего гипоталамуса
проецируются в старую кору.
98
Онтогенез.
Онтогенез. В начальной стадии формирования мозга человека передний мозг является
закругленным концом нервной трубки. На этом этапе конечный мозг представлен тонкой
ростральной стенкой переднего мозга. Затем эта стенка выпячивается в дорсолатеральном
направлении и образует два мозговых пузыря. Полости этих пузырей образуют боковые
желудочки. На ростральном конце мозговой трубки и III желудочка находится участок ткани,
соединяющий оба пузыря конечного мозга. В дальнейшем он станет конечной пластинкой.
Дно мешка конечного мозга утолщается очень быстро, это утолщение образует полосатое тело
(стадия 10-20 мм). В дальнейшем оно делится на хвостатое ядро, скорлупу, бледный шар и
миндалину. По мере роста полушарий базальные ядра смещаются медиально и примерно на 10-й
неделе развития (40 мм) сливаются с промежуточным мозгом.
Растущие аксоны покидают кору примерно на 8-й неделе развития (стадия зародыша — 23 мм).
Волокна, которые не заканчиваются в полосатом теле, идут вдоль плоскости слияния основания
конечного мозга с промежуточным, образуя внутреннюю капсулу. Волокна, идущие каудально и
огибающие снизу средний мозг, образуют ножки мозга. Затем на вентральной поверхности
продолговатого мозга они образуют пирамиды. Уходя в спинной мозг, пирамидные пути
перекрещиваются и оканчиваются на мотонейронах спинного мозга. Ассоциативные проводящие
пути начинают выявляться в конце второго месяца развития. Свод появляется из гиппокампа в
конце третьего месяца развития.
Мозолистое тело появляется в начале четвертого месяца развития. Оно начинает формироваться в
концевой пластинке в виде пучка поперечных волокон, лежащих над комиссурой гиппокампа.
Мозолистое тело растет очень быстро в каудальном направлении и смещает туда же комиссуру
гиппокампа и свод. Гиппокамп также смещается назад.
Пузыри конечного мозга разрастаются в ростральном, дорсальном и каудальном направлениях. На
ранних стадиях развития (до 3—4-го месяца) пузыри имеют очень тонкую стенку и утолщаются
медленно.
Борозды и извилины начинают формироваться только с 11 — 12-й недели. Первыми появляются
латеральная и гиппокампова извилины. Затем формирование борозд протекает очень быстро. К
моменту рождения существуют все основные извилины.
7. ОРГАНЫ ЧУВСТВ
В процессе эволюции у животных сформировались системы восприятия окружающего мира —
экстерорецепторы — и системы оценки состояния собственных систем организма —
интерорецепторы.
Анатомически органы чувств состоят из воспринимающей части — совокупности рецепторов,

проводящих нервных путей, промежуточных центров обработки информации и коркового отдела,
в котором происходит распознавание стимулов.
Разные по своему строению и внутренней организации рецепторы позволяют организму
воспринимать различные по своим свойствам внешние и внутренние сигналы. К ним относятся:
световые, звуковые, обонятельные, тактильные, вкусовые, температурные, механические, болевые
и другие сигналы или раздражители.
Рецепторный компонент органов чувств состоит из специальных клеток, имеющих специфическое
строение и способных воспринимать определенный тип сигнала. Проводниковый компонент
представлен нервными волокнами, идущими к соответствующим структурам центральной
нервной системы — спинному и головному мозгу, где и происходит обработка непрерывно
поступающей информации.
7.1. Зрительная система
Орган зрения состоит из глазного яблока, расположенного в глазнице, и зрительного нерва,
идущего к соответствующим участкам коры головного мозга
Ранее уже упоминалось, что в процессе эмбриогенеза архэнцефалона происходит превращение его
в передний мозговой пузырь, из дорсолатеральных стенок которого выступают два глазных
пузыря. В дальнейшем из них развиваются некоторые компоненты
100
Рис. 53. Схема горизонтального сечения правого глаза:
1 — хрусталик; 2 — зрительная ось; 3 — центральная ямка; 4 — желтое пятно; 5 — диск зрительного нерва;
6 — зрительный нерв; 7 — сетчатка; 8 — сосудистая оболочка; 9 — склера; 10 — стекловидное тело; 11 —
ресничная мышца; 12 — волокна пояска; 13 — задняя камера; 14 — передняя камера; 15 — роговица; 16 —
радужная оболочка; 17 — конъюнктива
зрительной системы, в том числе проводящие пути промежуточного мозга.
Глазное яблоко
Глазное яблоко имеет шаровидную форму, которая изменяется в процессе постнатального
развития новорожденного. Оно состоит из ядра, покрытого тремя оболочками — фиброзной,
сосудистой и сетчатой (внутренней) (рис. 53).
Фиброзная оболочка подразделяется на прозрачную переднюю часть — роговицу (рис. 53, 15) и
заднюю — склеру (рис. 53, 9). Склера (sclera) представляет собой плотную соединительную ткань,
образованную пучками коллагеновых волокон. Сзади на склере находится решетчатая пластинка,
через которую проходят волокна зрительного нерва. В толще склеры, на границе соединения ее с
роговицей, имеется сеть мелких полостей, образующих венозный синус склеры, через который
происходит отток жидкости из передней камеры глаза.
Роговица (cornea) — это выпуклая пластинка блюдцеобразной формы
Роговица (cornea) — это выпуклая пластинка блюдцеобразной формы, круглый край (лимб)
которой переходит в склеру. Толщина роговицы от 0,8 до 1, 1 мм. Роговица лишена кровеносных
сосудов, и ее питание происходит за счет лимфы.
Сосудистая оболочка глазного яблока находится под склерой и состоит из собственно сосудистой
оболочки, ресничного тела и радужки. Ресничное тело участвует в аккомодации глаза,
поддерживая, фиксируя и растягивая хрусталик. Большая часть реснич-
101
ного тела — это ресничная мышца (рис. 53, 11), образованная пучками миоцитов, среди которых
различают продольные, циркулярные и радиальные волокна.
Ресничное тело спереди продолжается в радужку (рис. 53, 16), которая представляет собой