Файл: Воронова, Климова, Менджерицкий Анатомия центральной нервной системы.pdf
Добавлен: 19.02.2019
Просмотров: 6656
Скачиваний: 32

удлиняются и превращаются в дендриты и аксон, которые на данном этапе лишены миелиновых
оболочек. Миелинизация начинается с пятого месяца пренатального развития и полностью
завершается лишь в возрасте 5—7 лет. На пятом же месяце появляются синапсы. Миелиновая
оболочка формируется в пределах ЦНС олигодендроцитами, а в периферической нервной системе
— Шванновскими клетками.
В процессе эмбрионального развития формируются отростки и у клеток макроглии (астроцитов и
олигодендроцитов). Клетки мик-
31
роглии образуются из мезенхимы и появляются в ЦНС вместе с прорастанием в нее кровеносных
сосудов.
Клетки ганглиозных валиков дифференцируются сначала в биполярные, а затем в
псевдоуниполярные чувствительные нервные клетки, центральный отросток которых уходит в
ЦНС, а периферический — к рецепторам других тканей и органов, образуя афферентную часть
периферической соматической нервной системы. Эфферентная часть нервной системы состоит из
аксонов мотонейронов вентральных отделов нервной трубки.
В первые месяцы постнатального онтогенеза продолжается интенсивный рост аксонов и
дендритов и резко возрастает количество синапсов в связи с развитием нейронных сетей.
Эмбриогенез головного мозга начинается с развития в передней (ростральной) части мозговой
трубки двух первичных мозговых пузырей, возникающих в результате неравномерного роста
стенок нервной трубки (архэнцефалон и дейтерэнцефалон). Дейтерэнцефалон, как и задняя часть
мозговой трубки (впоследствии спинной мозг), располагается над хордой. Архэнцефалон
закладывается впереди нее. Затем в начале четвертой недели у зародыша дейтерэнцефалон
делится на средний (mesencephalon) и ромбовидный (rhombencephalon) пузыри. А архэнцефалон
превращается на этой (трехпузырной) стадии в передний мозговой пузырь (prosencephalon) (рис.
17, 9). В нижней части переднего мозга выпячиваются обонятельные лопасти (из них развиваются
обонятельный эпителий носовой полости, обонятельные луковицы и тракты). Из
дорсолатеральных стенок переднего мозгового пузыря выступают два глазных пузыря. В
дальнейшем из них развиваются сетчатка глаз, зрительные нервы и тракты.
На шестой неделе эмбрионального развития передний и ромбовидный пузыри делятся каждый на
два и наступает пятипузырная стадия (рис. 17).
Передний пузырь — конечный мозг — разделяется продольной щелью на два полушария. Полость
также делится, образуя боковые желудочки. Мозговое вещество увеличивается неравномерно, и на
поверхности полушарий образуются многочисленные складки — извилины, отделенные друг от
друга более или менее глубокими бороздами и щелями (рис. 18). Каждое полушарие разделяется
на четыре доли, в соответствие с этим полости боковых желудочков делятся также на 4 части:
центральный отдел и три рога желудочка. Из мезенхимы, окружающей мозг зародыша,
развиваются оболочки мозга. Серое вещество располагается и на периферии, образуя кору
32

Рис. 18. Этапы развития головного мозга человека
больших полушарий, и в основании полушарий, образуя подкорковые ядра.
Задняя часть переднего пузыря остается неразделенной и называется теперь промежуточным
мозгом (рис. 17, 10). Функционально и морфологически он связан с органом зрения. На стадии,
когда границы с конечным мозгом слабо выражены, из базальной части боковых стенок
образуются парные выросты — глазные пузыри (рис. 17, 8), которые соединяются с местом их
происхождения при помощи глазных стебельков, впоследствии превращающихся в зрительные
нервы. Наибольшей толщины достигают боковые стенки промежуточного мозга, которые
преобразуются в зрительные бугры, или таламус. В соответствии с этим полость III желудочка
превращается в узкую сагиттальную щель. В вентральной области (гипоталамус) образуется
непарное выпячивание — воронка, из нижнего конца которой происходит задняя мозговая доля
гипофиза — нейрогипофиз.
33
Третий мозговой пузырь превращается в средний мозг (рис. 17, 5), который развивается наиболее
просто и отстает в росте. Стенки его утолщаются равномерно, а полость превращается в узкий
канал — Сильвиев водопровод, соединяющий III и IV желудочки. Из дорсальной стенки
развивается четверохолмие, а из вентральной — ножки среднего мозга.
Ромбовидный мозг делится на задний и добавочный. Из заднего формируется мозжечок (рис. 17,
12) — сначала червь мозжечка, а затем полушария, а также мост (рис. 17, 11). Добавочный мозг
превращается в продолговатый мозг. Стенки ромбовидного мозга утолщаются — как с боков, так
и на дне, только крыша остается в виде тончайшей пластинки. Полость превращается в IV
желудочек, который сообщается с Сильвиевым водопроводом и с центральным каналом спинного
мозга.
В результате неравномерного развития мозговых пузырей мозговая трубка начинает изгибаться
(на уровне среднего мозга — теменной прогиб, в области заднего мозга — мостовой и в месте
перехода добавочного мозга в спинной — затылочный прогиб). Теменной и затылочный прогибы
обращены наружу, а мостовой — внутрь (рис. 17; 18).
Структуры головного мозга, формирующиеся из первичного мозгового пузыря: средний, задний и
добавочный мозг — составляют ствол головного мозга (trùncus cerebri). Он является ростральным
продолжением спинного мозга и имеет с ним общие черты строения. Проходящая по латеральным
стенкам спинного мозга и стволового отдела головного мозга парная пограничная борозда (sulcus
limitons) делит мозговую трубку на основную (вентральную) и крыловидную (дорзальную)
пластинки. Из основной пластинки формируются моторные структуры (передние рога спинного
мозга, двигательные ядра черепно-мозговых нервов). Над пограничной бороздой из крыловидной
пластинки развиваются сенсорные структуры (задние рога спинного мозга, сенсорные ядра ствола
мозга), в пределах самой пограничной борозды — центры вегетативной нервной системы.


Производные архэнцефалона (telencephalon и diencéphalon) создают подкорковые структуры и
кору. Здесь нет основной пластинки (она заканчивается в среднем мозге), следовательно, и нет
двигательных и вегетативных ядер. Весь передний мозг развивается из крыловидной пластинки,
поэтому в нем имеются лишь сенсорные структуры (см. рис.18).
34
Постнатальный онтогенез нервной системы человека начинается с момента рождения ребенка.
Головной мозг новорожденного весит 300—400 г. Вскоре после рождения прекращается
образование из нейробластов новых нейронов, сами нейроны не делятся. Однако к восьмому
месяцу после рождения вес мозга удваивается, а к 4—5 годам утраивается. Масса мозга растет в
основном за счет увеличения количества отростков и их миелинизации. Максимального веса мозг
мужчин достигает к 20—29 годам, а женщин к 15—19. После 50 лет мозг уплощается, вес его
падает и в старости может уменьшиться на 100 г.
5. ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА
Вегетативная нервная система регулирует внутренние процессы, обеспечивающие жизнь
организма, такие как пищеварение, дыхание, сердечно-сосудистая деятельность.
Центральные структуры вегетативной нервной системы расположены в головном и спинном
мозге. В головном мозге это, прежде всего, гипоталамические центры, обеспечивающие
постоянство внутренней среды организма, а также стволовые вегетативные ядра. В спинном мозге
нейроны вегетативной нервной системы располагаются на границе между базальной и
крыловидной пластинами, образуя боковые рога серого вещества.
Периферические части вегетативной нервной системы состоят из ганглиев, представляющих собой
скопления нервных клеток, лежащих вне пределов ЦНС, и волокон. Эфферентные волокна
центральных структур вегетативной нервной системы выходят из ЦНС в составе смешанных
черепно-мозговых или по передним корешкам спинномозговых нервов. Затем покидают общий
нервный ствол и переключаются в ганглиях. Афферентные волокна заходят в ЦНС вместе с
чувствительными соматическими волокнами через задние корешки спинного мозга или в составе
черепно-мозговых нервов.
Ганглии, в зависимости от местоположения, делятся на: паравертебральные (лежащие в
непосредственной близости от позвоночника), превертебральные (удаленные от позвоночника),
экстрамуральные (лежащие поблизости от иннервируемого органа) и интрамуральные
(расположенные непосредственно в стенке органа). Волокна, подходящие к ганглию
(преганглионарные), покрыты миелином, волокна, покидающие ганглий (постганглионарные),
немиелинизированы и имеют серый цвет.
В вегетативных ганглиях находятся афферентные, эфферентные и ассоциативные нейроны. Для
этих ганглиев характерно, что
36
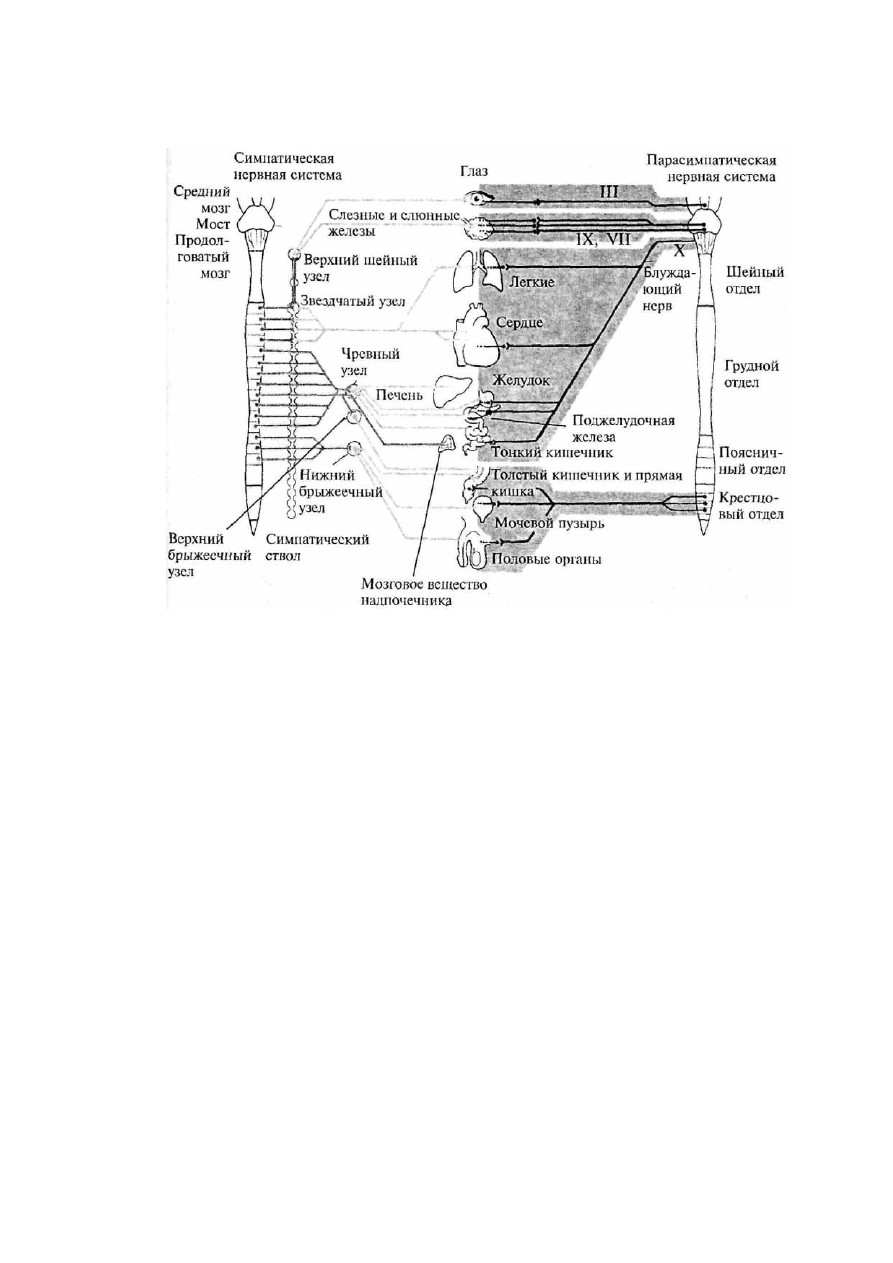
Рис. 19. Схема строения вегетативной нервной системы. Римскими цифрами
обозначены соответствующие пары черепно-мозговых нервов
количество
постганглионарных
волокон
гораздо
больше
количества
подходящих
преганглионарных. Эфферентные вегетативные волокна оканчиваются на гладкой мускулатуре
внутренних органов, сердечной мышце и железах.
Вся вегетативная нервная система состоит из двух отделов: парасимпатического и
симпатического. Оба эти отдела иннервируют одни и те же органы, т.е. к каждому вегетативному
органу подходят как парасимпатические, так и симпатические окончания, часто оказывая на него
противоположное действие (рис. 19).
5.1. Парасимпатический отдел вегетативной нервной системы
Центральные структуры
Центральные структуры парасимпатического отдела вегетативной нервной системы
расположены в стволе мозга (средний мозг,
37
Варолиев мост и продолговатый мозг) и в крестцовом отделе спинного мозга. Периферические
части образованы эктрамуральными и интрамуральными ганглиями и нервами.
Из среднего мозга парасимпатические ветви уходят в составе глазодвигательного нерва (III пара).
Затем преганглионарные волокна направляются к ресничному экстрамуральному ганглию
глазницы. Постганглионарные волокна этого ганглия иннервируют гладкую мускулатуру
ресничного тела и кольцевых мышц зрачка, т.е. являются двигательными.
Варолиев мост покидается парасимпатическими волокнами в составе лицевого нерва (VII пара).
На периферии они образуют преганглионарные веточки нескольких экстрамуральных узлов,
иннервирующих железы слизистой оболочки носа и нёба, слезные железы, подчелюстную и
подъязычную слюнные железы. Таким образом, парасимпатические веточки лицевого нерва
являются секреторными.
Из продолговатого мозга в составе языкоглоточного нерва (IX пара) уходят также секреторные
парасимпатическое ветви, которые направляются к ушному экстрамуральному ганглию,
иннервирующему околоушные слюнные железы и железы слизистой щек и губ.
Блуждающий нерв (X пара) является самой значительной частью парасимпатического отдела
вегетативной нервной системы. Его ветви направляются к сердечному, бронхиальному и чревному

сплетениям, а также к интрамуральным узлам в стенках внутренних органов грудной, брюшной
полостей и полости большого таза.
Парасимпатические ветви крестцовой части спинного мозга берут начало в боковых рогах серого
вещества второго—четвертого крестцовых сегментов и направляются к превертебральным
ганглиям нижнего подчревного сплетения и интрамуральным ганглиям органов малого таза (см.
рис. 19).
Окончаниями парасимпатического отдела вегетативной нервной системы выделяется медиатор
ацетилхолин.
Парасимпатический отдел вегетативной нервной системы регулирует работу внутренних органов
в условиях покоя. Его активация способствует снижению частоты и силы сердечных сокращений,
снижению кровяного давления, увеличению как двигательной, так и секреторной активности
пищеварительного тракта.
38
5.2. Симпатический отдел вегетативной нервной системы
Центральные структуры
Центральные структуры симпатического отдела вегетативной нервной системы расположены в
спинном мозге. Они занимают пространство боковых рогов серого вещества от восьмого шейного
сегмента
до
второго-третьего
поясничного
(спинномозговой
центр
Якобсона).
Миелинизированные аксоны этого центра выходят в составе передних корешков спинного мозга.
Периферическая часть
Периферическая часть симпатического отдела состоит из двух пограничных стволов — цепочек
паравертебральных ганглиев, лежащих по краям позвоночника. Ганглии в цепочке связаны между
собой межузловыми ветвями (коннективами). Существуют и комиссуральные связи между
симметричными ганглиями. В шейном и нижнем крестцовом отделах симпатического ствола
преганглионарные нервы подходят к ганглиям не из своих сегментов спинного мозга, а из ниже-
или вышележащих сегментов через коннективы ствола. В этом случае веточки проходят через
ганглии, не переключаясь в них и оставаясь миелинизированными.
В шейном отделе имеются три шейных узла, образовавшиеся в онтогенезе при слиянии восьми
симпатических ганглиев. Два верхних шейных узла иннервируют сонную артерию, глотку,
пищевод, слюнные и щитовидные железы и сердце. Нижний шейный узел, в свою очередь,
сливается с верхним грудным симпатическим узлом, образуя крупный звездчатый ганглий.
Звездчатый ганглий иннервирует позвоночную артерию, органы грудной полости (пищевод,
трахею, вилочковую железу, аорту) и сердечную мышцу.
В грудном отделе имеется 10—12 ганглиев. Постганглионарные ветви первых пяти из них
направляются к плевре, сердечному, легочному и аортальному сплетениям. Узлы с 6-го по 9-й
образуют большой чревный нерв, который, пройдя в брюшную полость, оканчивается в
превертебральных узлах самого крупного нервного сплетения брюшной полости — чревного или
солнечного сплетения. В его состав входят как симпатические, так и парасимпатические волокна.
От чревного сплетения радиально отходит множество нервов, образующих вторичные сплетения
(отсюда и название — солнечное сплетение). В узлах солнечного сплетения оканчиваются волокна
многих внутренностных нервов. От узлов этого сплетения берут начало постганглионарные нервы,
иннер-
39
вирующие почти все органы брюшной полости. Последние 2—3 узла грудного отдела своими
ветвями формируют малый чревный нерв, который также уходит к узлам солнечного сплетения.
В брюшной части пограничного симпатического ствола имеется 4—5 поясничных узла, от
которых идут ветви к брюшной части аорты, половым органам, брыжеечному ганглию,
кишечнику. В тазовой части симпатического ствола лежат 4 крестцовых узла и один непарный
копчиковый узел. Через тазовое сплетение они иннервируют тазовые органы.
Симпатические афферентные (чувствительные) волокна от внутренних органов, несущие
информацию от многочисленных интерорецепторов, идут в составе симпатических нервов и
вступают в спинной мозг по его задним корешкам, как и чувствительные соматические
афференты.
Окончания симпатических волокон выделяют в качестве медиатора норадренилин и адреналин.
Симпатический отдел вегетативной нервной системы увеличивает свою активность при
необходимости мобилизации ресурсов организма. Под действием импульсов, приходящих по