ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 22.02.2019
Просмотров: 15220
Скачиваний: 17

70
Розділ 2
Пригнічуючи виділення передньою часткою гіпофіза лютропіну, прогестерон
гальмує дозрівання вторинних фолікулів у яєчниках – овуляцію, а у молочних залозах
він викликає проліферацію епітелію у їх аценозній частині.
Концентрація прогестерону у крові починає зростати з 4-го дня циклу, досягаючи
максимуму на 7–8 день, а за деякими авторами на 10–12-й день, з 14–15-го дня вона
знижується і особливо різко перед наступною охотою.
Імпульс для секреції прогестерону (як і естрогенів) надходить з кори головно-
го мозку. Термін функціонування жовтих тіл контролюється маткою, у епітеліальних
клітинах якої синтезується біологічно активна речовина лютеолітичної дії – проста-
гландин F
2α
, який по кровоносних судинах доноситься до яєчника, акумулюється у
лютеальній тканині і викликає швидку її регресію.
Гормон жовтого тіла і його синтетичні замінники останнім часом широко застосо-
вуються для вкорочення сильно вираженої тічки, попередження спонтанних абортів,
синхронізації охоти і т. п.
Чоловічі статеві гормони (андрогени). Сюди належать андростерон, дигідро-
андростерон, тестостерон і споріднені з ними речовини. Вважають, що первинними
статевими гормонами є тестостерон, а андростерон і дигідроандростерон – продукти
біологічних змін його в організмі. У хімічному відношенні андрогени належать до тієї
ж групи стероїдних гормонів, що й естрогени. Вони викликають у молодих самців
розвиток вторинних статевих ознак і покручених каналиків сім’яника, а у статеводоз-
рілих тварин – статевий потяг і впливають на вираженість вторинних статевих рис.
При розладах регуляції статевої функції у самок андрогени можуть викликати в них
маскулінізацію.
Утворюються андрогени у сім’яниках та корі наднирників, піддаються в організмі
метаболічним перетворенням і виводяться з сечею у вигляді 17-кетостероїдів.
Тестостерон відіграє роль не лише у репродуктивній функції. Він стимулює також
синтез білка, чому самці мають більшу масу тіла, ніж самки. Крім того, тестостерон
може підсилювати еритропоез.
Схему регуляції статевої функції у самок можна уявити собі так: імпульси, що над-
ходять у кору головного мозку від зовнішнього середовища (через органи слуху, зору,
нюху, теплові та інші подразники), особливо сильні у присутності самця, передаються
по центробіжних нервах у відповідні центри кори головного мозку, сумуються тут
з імпульсами від внутрішніх органів, переробляються і передаються у гіпоталамус,
викликаючи в ньому стан підвищеної або ж зниженої функціональної активності. У
першому випадку в супраоптичних та паравентрикулярних ядрах гіпоталамуса виро-
бляється нейросекрет (рилізінг-фактор), який, поступаючи у гіпофіз, стимулює виді-
лення ним фолітропіну. Останній поступає у кров і, доносячись до яєчника, викликає
ріст і дозрівання фолікулів, що супроводжується синтезом у фолікулярних клітинах
естрогенів. Діючи на хеморецептори та аналізатори головного мозку, вони виклика-
ють у тварини тічку, охоту та загальне збудження (рис. 18).
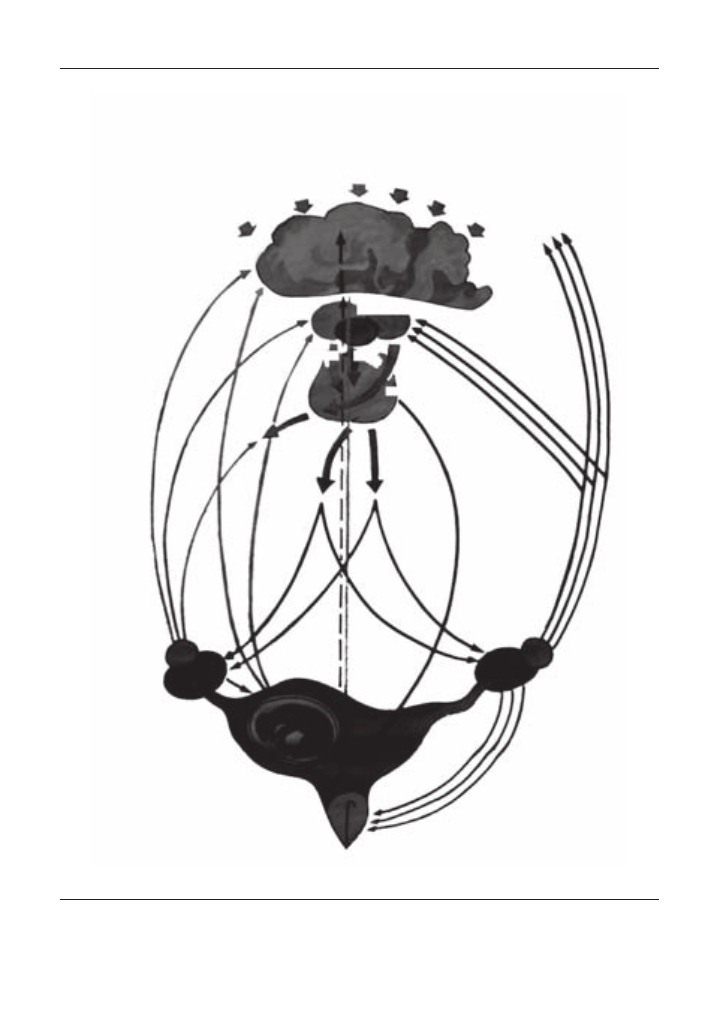
Фізіологічні особливості статевих органів свійських тварин
71
Рис. 18. Схема регуляції статевого циклу.
КО
Р
А
Г
О
Л
О
ВНО
ГО
МО
З
КУ
СТ. ОХОТА
ЕСТРОГЕНИ
фолікул
тічка
жовте тіло
гіпоталамус
гіпофіз
рф
гн
ФСГ
ЛГ
ЕСТРОГЕНИ
ЗОВНІШНІ ФАКТОРИ
ін
со
ля
ція
ре
це
пт
ори
ш
кір
и
і о
че
й
кор
мові
рец
ептор
и
тра
вного
трак
ту
самец
ь
відчутт
я
нюхові слухові з
оров
і та
кти
льн
і
Г
А
Л
Ь
М
У
В
А
Н
Н
Я
Т
ІЧ
К
И
Г
О
Р
М
О
Н
И
П
Л
А
Ц
Е
Н
Т
И
О
К
С
И
Т
О
Ц
И
Н
72
Розділ 2
Високі концентрації естрогенів у крові пригнічують виділення фолітропіну і сти-
мулюють виділення лютропіну, який викликає овуляцію і лютеїнізацію зернистої обо-
лонки фолікула. Жовте тіло, що виникло після овуляції, виділяє прогестерон, який
гальмує дальше виділення лютропіну, стимулює лютеотропну функцію і не перешко-
джає секреції фолітропіну, а у вагітних самок стимулює лютеотропну функцію гіпо-
фіза. На цьому фоні зростає секреція прогестерону.
При відсутності запліднення ендометрій виробляє в кінці статевого циклу люте-
олітичний фактор (ПГ-F
2α
), що викликає регресію жовтого тіла і статевий цикл по-
новлюється.
У великої рогатої худоби перша після овуляції хвиля росту фолікулів завершуєть-
ся їх атрезією, що стимулює нову хвилю росту фолікулів. Протягом статевого циклу
у них буває дві, а то й три хвилі росту фолікулів. Внаслідок останньої хвилі виникає
домінантний фолікул, який і овулює.
Для дозрівання фолікула, настання тічки та овуляції необхідне співвідношення
фолітропіну до лютропіну від 4 : 1 до 100 : 1.
Гормони яєчників, з одного боку, проявляють вплив на периферійні статеві орга-
ни, підготовляючи їх до імплантації ембріона, з другого боку, вони беруть участь у
регуляції функції системи гіпоталамус – гіпофіз за принципом зворотного зв’язку.
Підсумовуючи сказане, циклічний характер статевої функції самок можна
пояснити:
наявністю у центральній нервовій системі своєрідного біологічного годинни-
ка чи як ще його називають циклічного центру у вигляді гіпоталамусу;
періодичним виділенням цим центром рилізінг – гормонів;
відповідною зміною секреції гіпофізом фолітропіну та лютропіну;
послідовною зміною в яєчнику секреції естрогенів та прогестерону (фолікулі
нової та лютеїнової фаз статевого циклу);
наявністю системи зворотного зв’язку у ланцюгу гіпоталамус – гіпофіз – яєч-
ники та формуванням у корі головного мозку статевої домінанти, як ознаки
готовності організму до запліднення.
Якщо статевий цикл завершується заплідненням, то у корі головного мозку фор-
мується материнська домінанта, яка в кінці вагітності проявляється як родова, а зго-
дом – лактаційна домінанта.
Крім згаданих гормональних впливів, на функцію статевої системи впливають ті-
реотропний, адренокортикотропний, соматотропний, а також негіпофізарні гормони
епіфіза, наднирників, щитовидної та інших залоз.
Як відомо, під час вагітності роль ендокринної залози з багатогранною дією ви-
конує плацента. У її ворсинках виробляються значні кількості естрогенів та прогесте-
рону, а також гонадотропний гормон – сироватковий чи плацентарний (у жінок хорі-
альний) гонадотропін.
У кобил сироватковий гонадотропін з’являється у сироватці крові і лімфі з 4-го
дня жеребності, коли матка готується до імплантації зародка. Максимальної концен-

Фізіологічні особливості статевих органів свійських тварин
73
трації (80–160 МО/л) гонадотропіни досягають на 50–100-й день жеребності, опісля
концентрація їх швидко падає. У кобил сироватковий гонадотропін не проходить крізь
нирковий фільтр і тому затримується у крові, тоді як у жінок він проникає у сечу і на-
копичується там у великій кількості. Під час вагітності плацентарний гонадотропін
спочатку доповнює дію гіпофізарних гонадотропінів, а опісля заміняє їх.
Важливе значення для відтворної функції має також гормон задньої долі гіпофіза
окситоцин. Він стимулює скоротливу функцію матки і є однією з ланок механізму
родової діяльності. Істотна роль йому належить в регуляції лактації.
Ще у 60-х роках ХХ ст. було відмічено взаємозв’язок між станом матки та три-
валістю функціонування жовтого тіла. На цій підставі було зроблено висновок, що у
рогах матки виробляються лютеолітичні речовини місцевої дії. Дослідження останніх
років показали, що цю роль виконують простагландини (ПГ).
Вперше простагландини були відкриті в спермі. У 1930 р. американські гінекологи
Курцрок і Льоб повідомили, що матка жінки реагує на введення сперми скороченням
чи розслабленням. У 1935 р. шведський учений Ейлер довів, що тонізуючим впли-
вом на гладку мускулатуру володіє ліпідний екстракт із сперми і назвав його проста-
гландином (вважаючи, що він утворюється в простаті), а у 1957 р. простагландин F
2α
було екстраговано за допомогою фосфатного буфера з тканини
передміхурової залози
барана. При структурному аналізі простагландину в його білкових ланцюгах було ви-
явлено подвійний зв’язок та α-конфігурацію 9-ї гідроксильної групи. Ці особливості
було відображено в назві сполуки. За це відкриття С. Бергстрем був удостоєний Но-
белівської премії.
Нині відомо, що простагландини синтезуються у клітинних мембранах більшості
тканин ссавців.
Простагландин F
2α
володіє вираженою лютеолітичною дією, вкорочуючи наполо-
вину функцію жовтого тіла, що супроводжується зниженням у крові рівня прогесте-
рону та підвищенням концентрації стероїдів. Він також викликає початок родів та
швидке скорочення судин пупкового канатика.
Дослідження останніх років значно розширили розуміння суті статевого циклу та
його регуляції. Згідно П. К. Анохіна усі функціональні системи є саморегульованими,
а за К. В. Судаковим, зміна поведінки тварин визначається внутрішніми потребами
їх організму. За теорією функціональних систем основу статевого циклу складають
періодично повторювані циклічні зміни у системі гіпоталамус – гіпофіз – яєчники в
зв’язку з ростом та дозріванням фолікулів, овуляцією, формуванням і функціонуван-
ням жовтого тіла. Сюди також входять циклічні зміни матки, інших систем організму
та поведінки тварини.
Запорукою нормального прояву статевого циклу є синхронність дії основних ре-
гуляторних систем, що контролюють процес розмноження (А. Г. Нєжданов).
Гонадотропні гормони гіпофізу – фолітропін, що контролює ріст фолікулів та
синтез естрогенних гормонів, а також лютропін, що забезпечує дозрівання фолікулів,
74
Розділ 2
овуляцію, синтез прогестерону та естрогенів, проявляють свою активність лише при
певному їх співвідношенні.
З дозріванням у яєчниках фолікулів зростає виділення ними естрогенних гормо-
нів, які, в свою чергу, у передовуляційну фазу активують виділення гіпоталамусом
Гн–РГ, підвищують чутливість до нього аденогіпофізу, у якому різко зростає секреція
ЛГ, що забезпечує дозрівання і овуляцію фолікула.
З утворенням жовтого тіла і секрецією ним прогестерону за принципом зворот-
ного зв’язку гальмується дальше виділення ЛГ. Під впливом малої частоти виділення
Гн–РГ та ФСГ стимулюється ріст фолікулів. Поява домінантного фолікула супрово-
джується інтенсивною секрецією естрадіолу та інгібіну. Останній гальмує ріст інших
фолікулів. Така черговість проявляється декілька раз протягом статевого циклу, що
кожен раз завершується атрезією домінуючого фолікула, крім останнього, передову-
ляційного.
Стимулом частоти пульсації Гн–РГ, а за ним виділення ЛГ є зниження вмісту
прогестерону, а високі концентрації естрадіолу, навпаки, стимулюють виділення ЛГ
(Imakawa з співр.).
В регуляції статевої функції гіпофізу беруть також участь такі нейромедіатори
гіпоталамуса, як норадреналін, дофамін та серотонін, а також продуковані у ЦНС
опіоїдні пептиди енкефаліни, ендорфіни та динорфіни, зокрема у передачі нервових
імпульсів.
Формування феноменів стадії збудження статевого циклу відбувається під впли-
вом статевих гормонів лише при певному естрогено-прогестероновому та андрогено-
прогестероновому співвідношеннях (А. Г. Нєжданов). Перша хвиля росту фолікулів
у яєчниках у лютеїнову фазу статевого циклу не викликає змін у поведінці тварин.
Лише друга – третя хвиля росту фолікулів, на фоні регресії жовтого тіла, зниження
концентрації прогестерону, підвищення секреції андрогенів і естрогенів приводить до
формування стадії збудження статевого циклу та її феноменів. При цьому пік концен-
трації естрогенів припадає на період яскраво вираженої тічки та статевого збудження,
а тестостерону – на період прояву охоти.
Поряд з цим значну роль у регуляції статевої функції відводять тиреоїдним гормо-
нам, гормону епіфіза мелатоніну, пролактину і іншим біологічно-активним речовинам.
Штучна регуляція статевого циклу. Знання особливостей статевого циклу на-
штовхнуло вчених на ідею штучної регуляції відтворювальної функції, практичне за-
стосування таких заходів, як стимуляція багатоплідності, синхронізація охоти, віднов-
лення статевої циклічності, корекція часу овуляції, часу настання статевого сезону. У
1934 р. довів Лопирін А. І. можливість стимуляції суперовуляції, що опісля підтверди-
ли М. М. Завадовський, А. Л. Падучева, Роусон, Гердон, Еверілл, Робінсон та ін. Ши-
рокого практичного застосування при цьому набули сироватка крові жеребних кобил
(СЖК) та очищений її препарат (ГСЖК) – сироватковий гонадотропін, а також виго-
товлений з сечі жінок – хоріальний гонадотропін (ХГЛ). Вказані гонадотропіни володі-
ють фізіологічною дією ФСГ і ЛГ, причім в отриманій з 50-го по 70-й день жеребності