ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 22.02.2019
Просмотров: 15208
Скачиваний: 17
80
Розділ 2
екваційним, у результаті чого
сперматоцит 1-го порядку ді-
литься на два сперматоцити
2-го порядку, які в свою чергу
діляться на чотири спермати-
ди. Тобто з кожного сперма-
тоцита І порядку утворюється
чотири сперматиди.
Під час редукційного ме-
йотичного поділу ядра хромо-
соми об’єднуються попарно
і, не розщеплюючись вздовж,
розходяться до полюсів дочір-
них клітин – сперматоцитів
2-го порядку, забезпечуючи
їх половинним (гаплоїдним)
набором хромосом. Еквацій-
ний поділ (за типом мітозу)
відбувається дуже скоро після
редукційного, при цьому кож-
на хромосома розщеплюєть-
ся вздовж на дві половинки,
здатні відновити втрачену по-
ловину (редуплікація).
Сперматида, що при цьому
утворилась, містить зернис-
те ядро, у її цитоплазмі роз-
міщена центросома, оточена
оболонкою (центросферою чи
ідісомою), навколо якої згру-
пований у вигляді зерен чи
пластинок апарат Гольджі; хон-
дріосомний чи мітохондріаль-
ний апарат виступає у вигляді
дрібних паличок чи зерен.
Стадія формування від-
бувається у верхівках прото-
плазматичних відростків клі-
тин Сертолі, що виступають у
просвіт каналика, і характери-
зується складними морфоло-
гічними змінами сперматид.
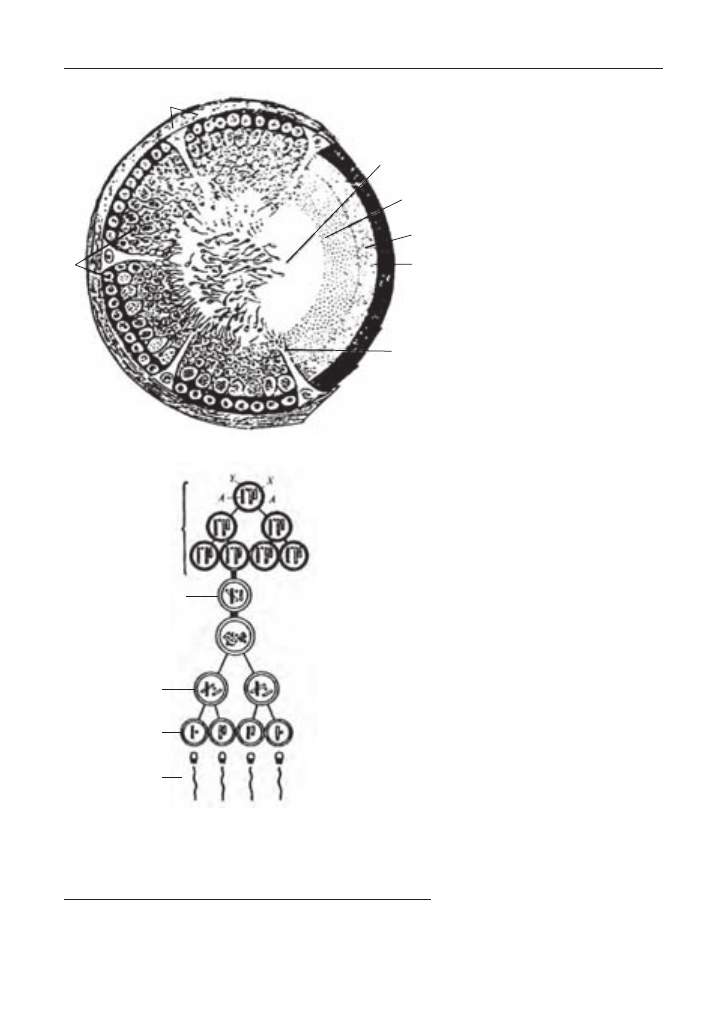
Рис. 19. Поперечний розріз сім’яного каналика (А) та
схема сперміогенезу (Б):
1 – сперматогенний епітелій; 2 – сполучно-тканинна стінка канальця;
3 – просвіт канальця зі сперміями; 4 – зона сперматид; 5 – зона сперма-
тоцитів; 6 – зона сперматогоній; 7 – синтицій Сертолі.
3
4
5
6
7
1
2
А
Б
1
3
6
9
10
стадія розмноження
стадія росту
стадія дозрівання
стадія формування

Фізіологічні особливості статевих органів свійських тварин
81
Клітинне ядро сперматиди переходить у верхню її частину і вона приймає овальну
форму. Розташовані біля переднього полюса ядра тільця Гольджі стають плоскими
і утворюють над ним ковпачок. Між полюсом і ковпачком виникає вакуоля з розмі-
щеною у ній ідіосомою (акросомною зерниною), яка переміщується до переднього
полюса ядра і разом з рідиною вакуолі утворює акросому. Спорожніла ідіосома разом
з апаратом Гольджі пересувається до протилежного полюса ядра і утворюють дві цен-
тросоми – проксимальну (нижчу), що наближається до ядра, і дистальну (дальшу),
розміщену на периферії сперматиди. Остання, давши початок вісьовий нитці джгутика
спермія, відокремлює від себе центросомне кільце навколо цієї нитки і перетворюєть-
ся з часом у основне кільце спермія, яке поступово пересувається разом з навколиш-
ньою протоплазмою у напрямку хвостика. Ділянка вісьової нитки джгутика, розта-
шована між центросомним і основним (базальним) кільцем, перетворюється згодом у
тіло спермія. З мітохондріального матеріалу цитоплазми формується спіральна нитка,
що обмотується навколо джгутика. Вона вкрита зверху протоплазматичним чохлом.
Плазма сперматид розподіляється переважно у голівці, шийці і тілі спермія і завершує
утворення сперміїв з характерною для них будовою – наявністю голівки, шийки, тіла
і хвостика. Сформовані спермії розріджують за допомогою ферменту гіалуронідази
протоплазматичний виріст клітини і виходять у просвіт каналика.
При розгляді під мікроскопом зрізу через звивистий каналик сім’яника можна по-
бачити, що сперміогенез має хвилеподібний характер: впродовж звивистого каналика
спостерігаються певні послідовні групи сперматогенного епітелію постійного клітин-
ного складу на різних стадіях дозрівання. Повна серія цитологічних змін між двома од-
наковими групами сперматогенних клітин у одній ділянці звивистого каналика назива-
ється циклом сперматогенного епітелію. Різні автори ділять цей цикл на 8–12 стадій.
Тривалість циклу сперматогенного епітелію (стадія дозрівання, від поділу спер-
матоцитів 2-го порядку до виходу сперміїв у просвіт сім’яних канатиків) складає у
барана 10,4 дня, кнура – 8, бугая – 13,51, людини – 16 днів. Тобто, найінтенсивніше
він перебігає у кнура.
Сформовані спермії поступають у просвіт покручених каналиків і переміщаються
по них у прямі каналики, сітку сім’яника, сперміовиносні каналики і нарешті – у ка-
нал придатка сім’яника.
Просуваються спермії по каналиках під тиском нових порцій сформованих гамет,
а також під впливом скорочення еластичних волокон стінок сім’яних каналиків.
Застосовуючи рентгенівське опромінювання та мічені попередники ДНК, вдалося
встановити тривалість часу між першим поділом сперматогоній і появою сформованих
сперміїв у придатку сім’яника, тобто “тривалість сперміогенезу” у різних видів тва-
рин. Вона складає у бугая 50–55 днів, барана – 40–53, кнура – 35–40, півня – 24–27.
По придатку сім’яника спермії просуваються рухами війок сперміовиносних про-
ток, перистальтичними скороченнями канала придатка, постійним виділенням секрету
та сперміїв сім’яними каналиками, а також за рахунок еякуляції. У барана спермії про-
ходять придаток за 10–14, а то і за 14–21 день, бугая – за 11, кнура – за 10–14 днів.

82
Розділ 2
При частому одержанні сперми від плідників проходження їх через придаток при-
скорюється і в еякуляті зростає кількість неповноцінних сперміїв, хоча навіть при
статевому виснаженні у придатку залишається великий запас сперміїв.
Добова спермопродукція у баранів складає біля 5 млрд, а у кнура – 12–15 млрд. Вста-
новлено тісний взаємозв’язок між масою сім’яників та їх репродуктивною здатністю.
Сперміогенез продовжується безперервно, але запаси сперміїв у придатку не пе-
ревищують певної межі, оскільки вони регулюються особливим біологічним механіз-
мом. Так, організм звільнюється від зайвих сперміїв шляхом ресорбції (фагоцитозу)
або ж виділення з сечею. У бугаїв під час статевого спокою виявляли щодоби у добо-
вій сечі від 125 до 3675 млн сперміїв, а при регулярному одержанні сперми – від 106
до 405 млн. У баранів ця цифра коливалась від окремих сперміїв і до 2477 млн.
Загальна довжина каналу придатка сім’яника складає у домашніх тварин від 30 до
86 м (у бугая – 30 м, кнура – 64, жеребця – 80 м). Тут міститься у барана від 100 до
220 млрд сперміїв, бугая – 50–75 млрд, кнура – 175–200 млрд.
У зв’язку з цим придаток сім’яника вважається своєрідним сперміосховищем. Тут
вони зберігають свою живучість і запліднюючу здатність до двох місяців.
У голівці придатка сім’яника відбувається остаточне дозрівання сперміїв (за 6–8
днів). Проходження сперміями довгого шляху від місця утворення до місця збері-
гання (а це біля 75 см довжини звивистого каналика, 14 см – по прямому каналику
і приблизно 500 м загальної довжини сітки сім’яника) супроводжується дальшими
змінами, що розпочалися ще з першого поділу сперматоцитів. Відбуваються не лише
їх кількісні, а й якісні зміни ядра та протоплазми клітини.
У придатку сім’яника спермії вкриваються колоїдним покривом, що захищає їх
від несприятливих впливів. Просуваючись по сім’явиносному каналу голівки при-
датка спермії вкриваються виділюваним епітелієм ліпопротеїдним секретом, який
переміщається поступово по ньому від голівки та шийки до хвоста. Кисла реакція
секрету придатка (концентрація водневих іонів тут у 10 разів вища, ніж у сім’яни-
ку), низький вміст цукрів, низький парціальний тиск кисню та високий тиск СО
2
, та
нижча на 3–4 ºС температура у порожнині калитки – обумовлюють перехід сперміїв
у анабіотичний стан, а багата мережа кровоносних та лімфатичних судин у придатку
забезпечує постачання сперміїв необхідною для цього стану енергією та видалення
токсичних продуктів обміну. Все це, поряд з іншими остаточно не з’ясованими фак-
торами, сприяє тривалому збереженню сперміїв.
У придатку сім’яника спермії набувають також від’ємного електричного заряду,
який обумовлений електронегативним характером колоїдів поверхні клітин. Цей од-
нойменний електричний заряд перешкоджає зіткненню і аглютинації сперміїв. При
зменшенні електричного заряду сили відштовхування слабнуть і можливе зіткнення
сперміїв і склеювання їх клейкою поверхнею. Такі спермії не можуть рухатися і при-
йняти участь у процесі запліднення.
Проходження сперміїв вздовж каналу придатка супроводжується не тільки змі-
нами їх фізіологічних властивостей, а й форми. Так життєздатність і запліднююча
здатність сперміїв, добутих з хвоста придатка, значно вища, ніж сперміїв з голівки
Фізіологічні особливості статевих органів свійських тварин
83
придатка. Крім того, спермії голівки придатка мають на проксимальному кінці шийки
так звану протоплазматичну краплю. З проходженням сперміїв крізь хвостову части-
ну придатка крапля просувається на тіло спермія, а тоді спадає зовсім.
Накопичені у придатку сім’яника спермії поступово виводяться під час статевого
акту чи одержання сперми на штучну вагіну. Якщо ж тривалий час плідник не вико-
ристовується, то спермії старіють, відмирають і видаляються частково з сечею, част-
ково піддаються резорбції, а на їх місце просуваються новосформовані.
Перебіг cперміогенезу, кількість та повноцінність утворених сперміїв, поряд з ге-
нетичними факторами, у значній мірі визначаються умовами утримання, годівлі та
догляду самців. Неповноцінна годівля, інтенсивна експлуатація, відсутність моціону,
ожиріння, як і різноманітні захворювання і в першу чергу захворювання статевих ор-
ганів негативно позначаються на сперміогенезі, кількості та якості виділюваних під
час еякуляції сперміїв, їх живучості та запліднюючій здатності. У кінцевому рахунку
вони можуть бути причиною неплідності.
2.3. Статевий інстинкт, статеві рефлекси та статевий акт
Статеве дозрівання самців супроводжується формуванням у них статевого ін-
стинкту, що складається з комплексу умовних та безумовних статевих рефлексів. За-
вдяки цим рефлексам відбувається парування самців та самок, що здійснюється через
статевий акт. Статевий акт чи парування (копуляція, коїтус) – складний фізіологічний
процес взаємодій двох різностатевих організмів, що супроводжується виділенням
самцем сперми у статеві шляхи самки (осіменіння).
Статевий акт складається з комплексу безумовних рефлексів (статевого потягу,
ерекції, обіймального, парувального, еякуляції), на які нашаровується ряд умовних
рефлексів, які можуть їх підсилювати чи, навпаки, гальмувати. Характер прояву ста-
тевого акту у значній мірі залежить від умов, у яких перебуває тварина, фізіологічно-
го стану її організму, активності статевих рефлексів, типів нервової діяльності самця,
самки і т. п. Перш за все, активність статевих рефлексів у значній мірі залежить від
рівня годівлі, утримання та експлуатації тварин, а також вироблених у процесі їх жит-
тя умовних рефлексів.
Згадані вище безумовні рефлекси є окремими ланками статевого інстинкту, закла-
деного у внутрішньоутробний період на базі успадкованих видових статевих ознак.
Вони проявляються під час статевого акту як у самця, так і у самки, правда по-своєму.
Рефлекс статевого потягу (зближення), або локомоторний рефлекс, проявляється
характерною для тварин реакцією – посиленням зорових та слухових відчуттів у при-
сутності тварин протилежної статі. Вони намагаються наблизитися одне до одного,
обнюхати, притулитися і здійснити статевий акт. Поведінка окремих самок з чітко
вираженим рефлексом зближення нагадує поведінку самця; вони плигають на інших
самок, наближаються до самця, ходять за ним.
Рефлекс ерекції у самців проявляється сильним наповненням кров’ю печеристих
тіл статевого члена, що надає йому пружності і сприяє введенню його у піхву сам-
84
Розділ 2
ки. Спочатку наповнюються кров’ю печеристі тіла стовбура прутня, а після введення
його у піхву – і голівки. Ерекція – це фізіологічний процес, під час якого через судини
прутня протікає крові у декілька разів більше, ніж під час статевого спокою. Внаслі-
док скорочення сіднично-печеристого та цибулинно-печеристого м’язів корінь прутня
притискається до сідничних кісток і відплив крові від нього затримується. Кров’яний
тиск у артерії прутня при цьому різко підвищується, до 3/5 тиску у аорті, тоді як під
час статевого спокою він буває удвічі нижчим.
Імпульс до ерекції, що зароджується в корі головного мозку під впливом зорових,
слухових, нюхових та тактильних відчуттів, передається у центр ерекції у крижовій
частині спинного мозку, а звідтіль – до статевих органів.
Ерекцію можна викликати і без участі центральної нервової системи – шляхом
механічного подразнення прутня, електричного подразнення крижових нервів і т. п.
У процесі використання плідників у них виробляються умовні рефлекси на час, місце
одержання сперми та ін.
У самок рефлекс ерекції проявляється посиленим припливом крові до статевих
органів, внаслідок чого виникає гіперемія слизових оболонок вульви, присінку, піхви
та шийки матки; печеристі тіла клітора та присінку наповнюються кров’ю, що обу-
мовлює зіяння статевої щілини.
Обіймальний рефлекс чітко виражений у самців, особливо у жеребців та кнурів.
Наблизившись до самки, вони стрибають на неї і фіксуються на ній передніми кінців-
ками. Важлива роль при цьому належить умовним рефлексам. Так, спочатку молоді
самці роблять садку не лише на самок в охоті, але й на самок не в охоті і навіть на
самців. Жеребці і кнури роблять садку на чучело, навіть коли воно мало подібне на
живу тварину, але з часом у них виробляється умовний рефлекс на самку в охоті, під-
ставну тварину, обстановку у манежі і т. п.
Прояв обіймального рефлексу, як і всіх інших, залежить від типу вищої нервової ді-
яльності тварини і узгодження ланок її нейрогуморальної регуляції. У несприятливій для
спаровування обстановці, при поганих погодних умовах, а також при неповноцінній годів-
лі, утриманні та експлуатації, при розладах обміну речовин рефлекс може гальмуватися.
У самок обіймальний рефлекс проявляється непротивленням статевому акту, на-
маганню самця зробити садку. Вона допускає його до себе і наче зосереджується на
статевому акті.
Парувальний рефлекс у самців проявляється штовхальними рухами крупа, що
забезпечують сприйняття рецепторами шкіри прутня термічних та механічних по-
дразнень, які виникають під час його тертя об слизову оболонку піхви.
У самок парувальний рефлекс проявляється характерною позою, яка є найсприят-
ливішою для тісного контакту її геніталій із статевим членом самця. Самка при цьому
підіймає хвіст, відводить його набік, вигинає спину, у неї періодично скорочується
мускулатура вульви, піхви та матки, окремих груп м’язів тулуба.
Рефлекс еякуляції проявляється у самців скороченням м’язів статевого апарату
(придатка, сперміопроводів, сечівника та додаткових статевих залоз), які забезпечу-
ють виведення сперміїв і секретів додаткових залоз із статевої системи самця.