ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 22.02.2019
Просмотров: 15270
Скачиваний: 17

210
Розділ 7
7.4. Синхронізація охоти
Ефективність трансплантації ембріонів у значній мірі визначається синхронністю
статевої функції у донорів і реципієнтів. Суть синхронізації охоти полягає у застосу-
ванні окремих прийомів та біологічно активних препаратів для регулювання статевої
функції у телиць чи корів з метою викликання у них одночасної охоти в заздалегідь
намічений час.
У великих стадах щоденно буває певна кількість тварин в стані охоти, проте важ-
ко передбачити, у скількох з них і коли вона наступить. Тому, в практичних умовах
звичайно вдаються до штучної синхронізації охоти різними методами. Наприклад, в
практичній ветеринарії давно застосовується енуклеація (вилущування) жовтого тіла.
Приблизно через 4 дні у тварин відновлюється статева циклічність. Проте енуклеація
жовтого тіла може ускладнюватися травмами тканин яєчника, утворенням спайок і
навіть загибеллю тварини від крововиливу. Тому ширше застосовування знаходять
інші методи, в основі яких лежить стимулювання та пролонгація функції жовтого тіла
чи, навпаки, пригнічення її.
В першому випадку застосовують введення прогестерону чи його синтетичних
аналогів-прогестагенів (хлормадінонацетат, КАП; медроксипрогестерон – МПА, ме-
ленгестролацетат – МГА, мегестролацетат, амол, діамол, ІСІ-79939, ІСІ-80996 та ін.),
які блокують гонадотропну функцію гіпофізу, не порушуючи синтезу ФСГ та ЛГ, під-
тримуючи існування жовтого тіла та високий рівень прогестерону в крові. Фолікули
при цьому дозрівають лише до другої стадії, але тічка, охота та овуляція гальмуються.
Припинення введення тваринам прогестерону чи прогестагенів супроводжується
інтенсивним виділенням гіпофізарних гормонів і появою у них статевого збудження.
Правда, час настання тічки, охоти та овуляції у тварин широко варіює, від 2 до 6 днів,
знижується і заплідненість яйцеклітин. Тому в практиці трансплантації ембріонів цей
метод не знайшов застосування, його витіс-
нили простагландини.
Увага до простагландинів типу ПГ-F
2α
значно зросла після того, як було виявлено,
що синтез та обмін їх у матці, матковій вені
та яєчниковій артерії в кінці лютеальної фази
статевого циклу у овець та корів супроводжу-
ється різким зниженням концентрації про-
гестерону в корові. З’ясувалося, що ПГ-F
2α
є єдиним лютеолітичним фактором, що син-
тезується в матці і викликає регресію жовто-
го тіла і контролює таким чином тривалість
статевого циклу. Це не могло не привернути
до себе уваги і незабаром з’явився ряд син-
тетичних аналогів ПГ-F
2α
, таких як простин
(США), клопростенол, еструмат (Велико-
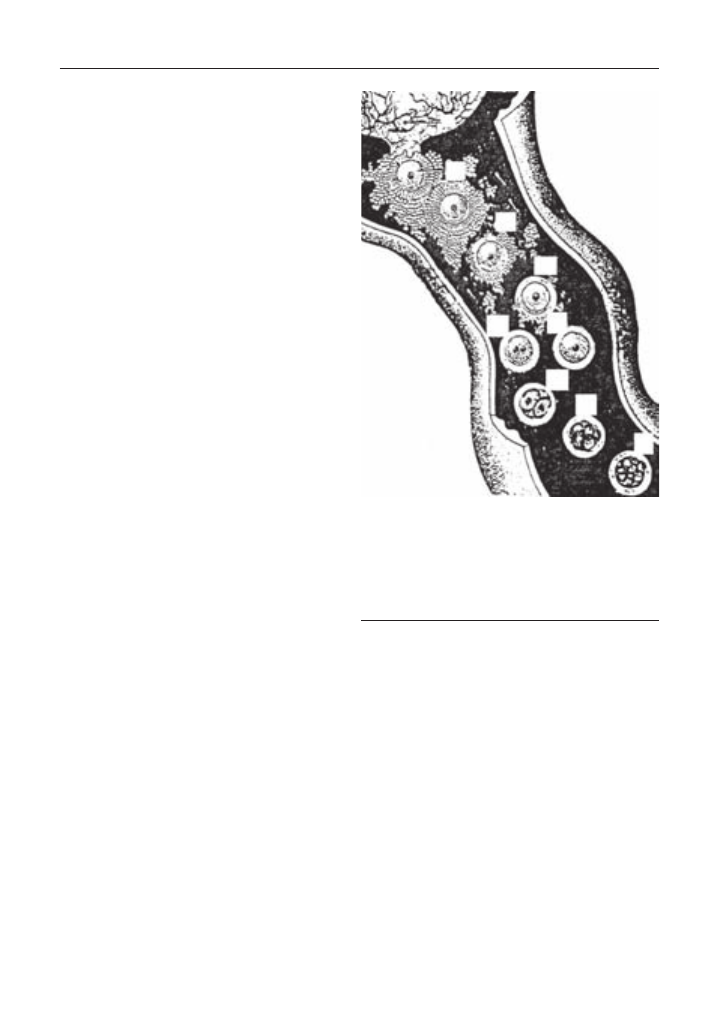
Рис. 42. Реакція яєчника на гормональ-
ну обробку.

Трансплантація ембріонів
211
британія), естрофан (Чехословаччина), ензапрост (Угорщина), естуфалан (СРСР) та
ін., які володіють значно вищою активністю, ніж природний ПГ-F
2α
.
При одноразовій обробці тварин лютеолітичною дозою ПГ-F
2α
приблизно у 66 %
з них поновлюється статева циклічність. Синхронізуючий ефект значно підвищуєть-
ся, якщо попередньо дослідити тварин ректально і залишити в групі лише особин з
активним жовтим тілом.
Для підвищення синхронізуючого ефекту ПГ-F
2α
його вводять двічі, з інтервалом
10–12 днів. Після повторної ін’єкції у 90–95 % тварин з’являється синхронна охота.
Приблизно у двох третин оброблених ПГ-F
2α
тварин з’являється охота через 38–
42 години, біля 10 % – на другу та четверту добу і лише у незначної частини – на
п’яту добу.
При досягненні повної синхронності статевої циклічності у донорів та реципієн-
тів Роусон з співробітниками домоглися вагітності реципієнтів у 91,1 % випадків, а
при появі її у реципієнтів на один день раніше чи пізніше, ніж у донорів, вагітність
наступала в 56,5 і 62,2 % випадків. Дещо нижчі, але подібні наслідки, отримали Ші
з співробітниками в досліді на 1 126 реципієнтах. Тобто, максимальне відхилення в
часі настання охоти між донорами та реципієнтами повинно бути плюс-мінус 1 день.
7.5. Осіменіння корів-донорів,
запліднення та ранні стадії розвитку ембріонів
У корів, оброблених гонадотропінами на 15–17-й день циклу, охота наступає через
3–5 днів, а при комбінованій стимуляції гонадотропінами та ПГ-F
2α
в лютеїнову ста-
дію циклу – через 75 годин після введення простагландину. При появі у тварин охоти
їх осіменяють.
Приблизно у 10 % донорів статевий цикл буває алібідним, хоча овуляція
відбувається.
Осіменяють донорів цервікально, з ректальною фіксацією шийки матки, з вико-
ристанням стерильних одноразових інструментів. У кожній дозі сперми повинно бути
не менше 50–100 млн рухливих сперміїв.
Оскільки охота та овуляція у оброблених гонадотропінами корів розтягується до
36–48 годин, то їх осіменяють 3–4 рази з інтервалами по 10–12 годин.
Оброблені гонадотропінами тварини відрізняються підвищеною чутливістю ста-
тевих органів до ректальної пальпації та інфекції, тому осіменяти їх слід обережно.
Необережні маніпуляції можуть викликати зміщення бахромки яйцепроводу, тоді не
всі яйцеклітини попадають в його просвіт. Крім того, необережна пальпація яєчника
може викликати передчасні розриви передовуляційних фолікулів і вихід недозрілих
яйцеклітин.
Введені в цервікальний канал спермії звичайно зберігають тут свою живучість до
двох діб, проте під впливом гормональних обробок донорів змінюється якість церві-
кального секрету, а отже і живучість, і запліднююча здатність сперміїв, що потрібно

212
Розділ 7
мати на увазі при осіменінні донорів. Просуваючись поступово в напрямку яйцепро-
водів спермії піддаються капацитації та селекції найбільш життєздатних.
Яйцеклітини, що попали після овуляції у яйцепровід, рухаються разом з течією
рідини в напрямку рога матки. У верхній третині яйцепроводу вони зустрічаються
зі сперміями і при сприятливих умовах переважна більшість з них запліднюється.
Запліднена яйцеклітина містить подвійний набір хромосом, тому вона називається
зиготою. Лімітуючим фактором відповідних змін сперміїв, яйцеклітин і самого за-
пліднення є середовище яйцепроводу.
Процес дальшого розвитку зиготи полягає перш за все в поділі її ядра, спочатку
на дві половинки (два бластомери), тоді на чотири, вісім і т. д., перетворюючись спо-
чатку в ембріони, а тоді в плоди.
Програма розвитку ембріона закодована в його ДНК. Послідовність та специфіч-
ність всіх його змін залежить від часу зняття блокади окремих детермінант, що були
раніше в неактивному стані. Кожна нова структура ембріона несе на своїй поверхні
нову антигенну інформацію, від якої може залежати гальмування одних та стимуляція
інших ділянок.
Приблизно через кожних 24 години кількість бластомерів у ембріоні подвоюється.
Прозора та жовткова оболонки при цьому деякий час зберігаються, загальний діаметр
ембріона залишається таким же, проте кожні наступні бластомери виявляються вдвічі
меншими попередніх, чому цей їх поділ ще називають дробленням.
Ранні ембріони затримуються 3–4 дні в яйцепроводах, цьому сприяє наявність між
яйцепроводом та рогом матки перешийка, що виконує роль адренергічного сфінктера.
В постовуляційну стадію під впливом прогестерону та ПГЕ сфінктер розслаблюється.
За час перебування ембріонів у яйцепроводі матка за допомогою фагоцитозу звіль-
нюється від залишків сперми, бактерій і т. п., в ендометрії відбуваються проліфера-
тивні зміни, підвищується активність маткових залоз, формується гістіотроф.
На 3–4-й день ембріон переміщається з яйцепроводу у матку – у великої рога-
тої худоби на стадії 8–16-ти бластомерів; у свиней – на стадії 4-х бластомерів; ко-
бил на стадії бластоцисти. Бластомери в цей час тісно прилягають один до одного
у вигляді шовковиці чи тутової ягоди, тому їх називають (ранньою) морулою. Кіль-
кість бластомерів збільшується і на 4–5-ту добу – це морула (16–32 бластомери), а на
6-ту–7-му – пізня морула (32–90 бластомерів).
На ранніх стадіях розвитку ембріона всі бластомери знаходяться в однакових умо-
вах, проте з часом вони диференціюються стосовно до їх локалізації, дещо змінюєть-
ся темп їх дроблення. Із зовнішнього шару бластомерів, що діляться дещо швидше,
формується живильний шар – трофобласт, що дасть початок судинній оболонці, а з
розміщеного в центрі грудки бластомерів, що діляться повільніше, – ембріобласт, з
якого з часом – розів’ється власне зародок (ембріон, а згодом плід) з водною та сечо-
вою оболонками.
На 7–8-му добу між трофобластом та ембріобластом з’являється щілина, наповне-
на рідиною – бластоцель, навколо якої перегруповуються бластомери: більші з них
(ембріобласт) зосереджуються на одному полюсі, а дрібніші – на протилежному. Ран-
Трансплантація ембріонів
213
ня бластоциста містить від 90 до 120-ти
бластомерів, поверхня клітинної маси
гладенька, рівномірна, товщина прозо-
рої оболонки – біля 12 мк.
В наступні дні відбувається даль-
ше збільшення кількості бластоме-
рів до 300–480 на стадії бластоцисти і
1 200–1 500 на стадії пізньої бластоцис-
ти. Дещо збільшується їх діаметр (до
140–200 мк), тоншає прозора оболонка,
розширюється порожнина бластоцисти,
вона займає всю прозору оболонку і на-
решті на 9–11-ту добу прозора оболонка
лопається (денудація) і ембріон “вилу-
плюється”. У овець це буває на 7–8-му,
свиней – на 6-ту добу розвитку ембрі-
она. Незабаром після денудації зарод-
ка відбувається обширна проліферація
трофобласта і помітний ріст в довжину
бластоцисти. Такий зародок може три-
валий час вільно переміщуватися в про-
світі матки, змінюючи поступово свій
розмір і форму. З 16–17-го дня у великої
рогатої худоби розпочинається імплан-
тація зародка.
Отже, найбільш придатним для ви-
мивання періодом розвитку ембріона є
7–8-й день, коли він ще захищений про-
зорою оболонкою.
7.6. Методи видобування ембріонів
Ефективність трансплантації ембріонів у значній мірі залежить від досконалості
застосованого методу видобування їх з геніталій донора. Із сказаного вище можна
зробити висновок, що на 4–5-ту добу основна кількість ембріонів у великої рогатої
худоби попадає в ріг матки. Проте суперовуляція значно розтягується в часі і не всі
ембріони одночасно з’являються у розі матки, частина з них може затримуватися в
яйцепроводі до 8-го дня. Поступово ембріони переміщаються від верхівки рогу матки
до його середньої і передньої частини.
Змінюється з часом і життєздатність ембріонів. У яйцепроводі практично всі вони
бувають нормальними, проте в міру переміщення їх у матку і збільшення строку пе-
Рис. 43. Запліднення і ранні стадії розвитку
ембріона у яйцепроводі самки:
1 – фолікул, що овулював; 2 – яйцеклітина, оточена про-
меневим вінцем; 3 – розсіювання променевого вінця;
4 – проникнення спермія крізь прозору оболонку; 5 – ста-
дія двох пронуклеусів; 6 – зигота; 7–9 – стадії двох, чоти-
рьох та восьми бластомерів.
1
2
3
4
5
6
7
8
9

214
Розділ 7
ребування їх в умовах
маткового середови-
ща (що не повністю
відповідає віку усіх
ембріонів),
зростає
кількість дегенерова-
них ембріонів.
До
середини
70-х років ембріони
у корів видобували
головно хірургічним
методом, вимиваючи
їх з яйцепроводу чи
рогу матки, залежно
від часу вимивання.
При цьому користу-
валися одним з трьох
до ступів: розрізом
верхнього склепіння піхви з використанням спеціального приладу (ефеменатора) або
без нього; лапаротомією по білій лінії живота із застосуванням наркозу; лапаротомією
з боковим розрізом в ділянці голодної ямки під місцевою анестезією.
Із запропонованих для хірургічного видобування ембріонів інструментів найпри-
датнішим виявився прилад Роуса і Даулінга, основою якого є гумовий катетер з на-
дувною кулькою в кінцевій частині, привідним та відвідним каналом.
Згодом запропоновано досконаліші металеві, гнучкі, пластмасові та гумові катетери.
Найширшого розповсюдження набув двоканальний урологічний катетер Фолея, в про-
світ якого перед застосуванням встановлюють гнучкий металевий стилет (рис. 44).
Для вимивання ембріонів користуються розчинами, які подібні за своїм складом
до секрету матки (рідина Дюльбекко, середовища Ігла, Паркера, ТМС-199, Хема-10,
Протасова, Менезо та ін.). Найбільш розповсюдженим є фосфатно-буферне сере-
довище (ФБС) Дюльбекко, до 1 л якого перед застосуванням додають 40 г бичого
альбуміну, 1,0 г глюкози, 0,036 г пірувату натрію та 100 тис. ОД пеніциліну (калієва
сіль). Осмотичний тиск розчинів повинен бути 300 міліосмоль, рН 7,2–7,6.
Хірургічне видобування ембріонів з рогів матки шляхом лапаротомії по білій
лінії. Витримують тварину два дні на голодній дієті, вводять їй внутрішньом’язово
для заспокоєння ромпун чи комбелен. Фіксують на операційному столі в спинному
положенні і проводять операцію під загальним наркозом. Підготувавши операційне
поле, розрізають черевну стінку по білій лінії довжиною 15–20 см спереду молочної
залози, виводять назовні один ріг матки з яйцепроводом і яєчником і підраховують в
ньому кількість жовтих тіл. Фіксують тіло матки в раневому отворі, проколюють стін-
ку рога матки біля біфуркації гострим гемостатичним пінцетом і вводять через утво-
Рис. 44. Гумовий катетер для нехірургічного вимивання
ембріонів:
1 – гумова трубка; 2 – надувна кулька; 3 – отвори для введення та виведення рідини;
4 – гумова “заглушка”; 5 – сталевий стилет; 6, 7 – надувний канал; 8 – розширення
каналу; 9 – зовнішній кінець катетера із замковим вузлом (10).
1
6
2
3
5
4
7
8
9
10
9
10
8
5