ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 22.11.2023
Просмотров: 554
Скачиваний: 3
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
;
■ наличие многоклеточных органов размножения — полового (гаметангий) и бесполого (спорангий). Мужские гаметангии высших растений называются антеридиями, женские — архегониями. Гаметангии высших растений (в отличие от низших) защищены оболочками из стерильных (бесплодных) клеток и (у отдельных групп растений) могут быть редуцированы, т.е. уменьшены и упрощены;
■ превращение зиготы в типичный многоклеточный зародыш, клетки которого первоначально не дифференцированы, но генетически детерминированы на специализацию в определенном направлении;
■ правильное чередование двух поколений — гаплоидного полового (гаметофит), развивающегося из споры, и диплоидного бесполого (спорофит), развивающегося из зиготы;
■ доминирование в жизненном цикле спорофита (у всех отделов кроме Моховидных);
■ разделение тела спорофита (у большинства отделов высших растений) на специализированные вегетативные органы — корень, стебель и листья.
Отделы:
Риниофиты (полностью исчезли)
Моховидные (25 000 видов)
Плауновидные (1 200 видов)
Хвощевидные (30 видов)
Папоротниковидные (10 000 видов)
Голосеменные (около 700 видов)
Покрытосеменные, или Цветковые (250 000 видов;)
Отличия высших растений от водорослей
Высшие растения, вероятно, произошли от каких-либо водорослей. Об этом свидетельствует то, что в геологической истории растительного мира высшим растениям предшествовали водоросли. В пользу этого предположения свидетельствуют и следующие факты: сходство наиболее древней вымершей группы высших растений - риниофитов – с водорослями, очень сходный характер их ветвления; сходство в чередовании поколений высших растений и многих водорослей; наличие жгутиков и способность к самостоятельному плаванию у
мужских половых клеток многих высших растений; сходство в строении и функции хлоропластов.
Предполагают, что высшие растения произошли скорее всего от зеленых водорослей, пресноводных или солоноватоводных. Они имели многоклеточные гаметангии, изоморфное чередование поколений в цикле развития.
Первыми наземными растениями, найденными в ископаемом состоянии, были риниофиты (риния, хорнея, хорнеофитон, спорогонитес, псилофит и др.).
После выхода на сушу высшие растения развивались в двух основных направлениях и образовали две большие эволюционные ветви – гаплоидную и диплоидную.
По имеющимся палеонтологическим данным, это произошло примерно 440 млн лет назад (силурийский период палеозойской эры). Известна точка зрения о том, что некие водоросли, жившие вблизи берега, сначала «высунули голову на воздух», затем заселили приливно-отливную зону, а потом, постепенно превращаясь в высшие растения, целиком вышли на берег. Впоследствии осуществилось постепенное завоевание ими суши.
С выходом растений на сушу у них появились ткани. В связи с тем, что воздух не может служить надежной опорой, у растений возникли механические ткани (древесные и лубяные волокна). Широкий диапазон изменения климатических факторов стал причиной формирования плотных покровных тканей (перидермы и корки). Нахождение воды и питательных веществ в почве обусловило появление проводящих тканей (ксилемы и флоэмы), формирование корня, стебля и листьев. Благодаря подвижности воздуха (ветру) у растений сформировались приспособления к опылению, распространению спор, пыльцы, плодов и семян.
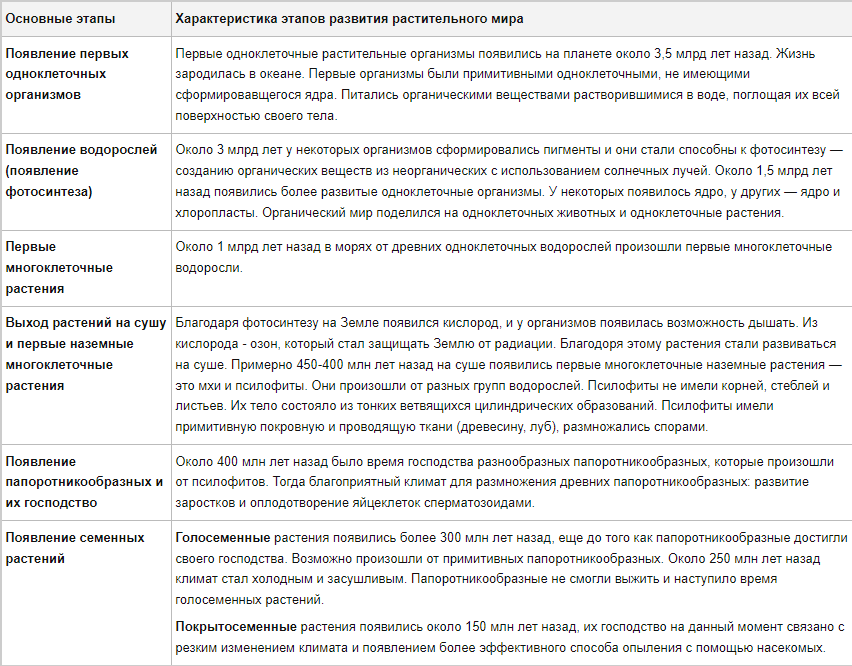
Палеозой:
В первых двух периодах эры палеозоя растительный мир был представлен преимущественно водорослями. В период силура появляются первые споровые растения, а в начале делура уже имеется множество простейших растений – риниофитов. К середине этого периода растительность развивается. Появились первые плауновые, прапапоротники, членистостебельные, прогимноспермы, голосеменные. Развивается покров почвы. Карбон ознаменовал появление хвощеподобных, древовидных плацновых, папоротников и папоротникообразных, кордаитов. Флора карбона со временем образовала толстый слой каменного угля, который добывается по сей день.
Мезозой:
С окончанием палеозоя вымирают папоротники, плауны и древесные хвощи. На смену им в триасовом периоде приходят хвойные и другие голосеменные растения. В юрском периоде вымирают уже голосеменные папоротники и появляются древесные покрытосеменные растения. Всю сушу покрывает обильная растительность, появляются предшественники сосен, кипарисов, мамонтовых деревьев. В меловом периоде развились первые растения с цветами. У них был тесный контакт с насекомыми, одни без других, по сути, не существовали. Поэтому за короткое время они распространились по всем уголкам планеты.
Палеозой:
На высоких широтах начинают преобладать покрытосеменные растения и хвойные. Зона экватора была покрыта дождевыми влажными лесами (пальмы, сандал, фикусы). В глубине материковых зон были распространены саванны и редкие леса. На средних широтах росли растения тропического типа – хлебные деревья, древовидные папоротники, банановые деревья, сандал.
Заполярье было покрыто широколиственными и хвойными деревьями. В неогене начинает развиваться флора современного Средиземного моря. На севере уже почти не было вечнозеленых растений. Выделяются таежная, тундровая и лесостепная зона. На месте саванн появляются пустыни или полупустыни.
РИНИОФИТЫ, первые наземные растения. Существовали в силуре – верхнем девоне. Примитивные сосудистые растения выс. 20–70 см, иногда до 3 м, без корней и листьев, с протостелическим (единый центральный проводящий пучок ксилемы, окружённый кольцом флоэмы) стеблем, преимущественно с дихотомическим вильчатым ветвлением и спорангиями на концах побегов, реже – вдоль побегов. Возможно, что спорофит и гаметофит существовали в виде самостоятельных растений. Росли по берегам водоёмов, располагаясь частично в воде или на насыщенных водой участках суши. Происхождение не ясно. Считаются предками других высших растений. Выход риниофитов на сушу – одно из важнейших событий в эволюции биосферы.
Риниофиты характеризовались верхушечным (терминальным) расположением спорангиев, отсутствием корней и листьев, дихотомическим, или дихоподиальным (псевдомоноподиальным), ветвлением спорофита, очень примитивным анатомическим строением и равноспоровостью. Проводящая система была слабо развита и представляла собой типичную протостелу. При развитии ксилемы первыми формировались трахеиды в центре ксилемы, т. е. ксилема была центрархная.
Филогенетическое значение: Риниофиты сам древн гр раст дала начало трем ветвям плауновидные, хвощевидные, папоротниковидные. От папоротников берут начало голосеменные, а от них цветковые. Благодаря им возникла теломная или эволюционная морфология, кот заним-ся изучением вегетативных органов высш раст.
Типы органов высших растений:
■ вегетативные (корень, стебель, лист);
■ генеративные (спорангии, антеридии, архегонии, цветок).
❖ Вегетативные органы — органы, выполняющие основные функции питания и обмена веществ с внешней средой, обеспечивая фотосинтез, водоснабжение, дыхание и рост растения.
Особенности вегетативных органов. Эти органы:
■ образуют систему побегов и корневую систему;
■ не участвуют в спорообразовании и половом размножении;
■ могут выполнять функцию вегетативного размножения;
■ при изменении функций претерпевают метаморфозы (видоизменяются).
❖ Генеративные (или репродуктивные) органы — специальные органы, обеспечивающие размножение растений. Генеративные органы бесполого размножения растений:
■ у споровых растений — спорангии,
■ у семенных растений — пыльцевой мешок и семязачаток.
Генеративные органы полового размножения растений:
■ мхов и папоротников — антеридии и архегонии,
■ цветковых растений — цветки, плоды, семена.
Первые достоверные наземные растения, известные только по спорам, датируются началом силурийского периода. Из верхнесилурийских и нижнедевонских отложений по сохранившимся макроостаткам или по отпечаткам органов описаны наземные растения. Эти первые известные нам высшие растения объединены в группу риниофитов. Несмотря на анатомическую и морфологическую простоту строения, это были уже типичные наземные растения. Об этом свидетельствует наличие кутинизированной эпидермы с устьицами, развитой водопроводящей системы, состоящей из трахеид, и наличие многоклеточных спорангиев с кутинизированными спорами. Следовательно, можно предположить, что процесс освоения суши растениями начался значительно раньше - в кембрии или в ордовике. В ходе длительного эволюционного процесса возникали новые виды, постепенно формировавшие типичные наземные растения
Исторически вегетативные органы высших растений возникли несколько позднее, чем ткани. Как указывалось ранее, тканевая дифференциация таллома началась при освоении суши, как следствие перехода растений к постоянной жизни в двух средах - воздушной и почвенной. Ткани имелись уже у древнейших наземных растений риниофитов. Однако само по себе вегетативное тело риниофитов представляло более или менее разветвленную безлистную ось. Отдельные цилиндрические веточки этой оси получили название теломов. Считается, что из этих веточек-теломов возникает побег. У древнейших высших растений побег и его видоизменения выполняли все основные функции вегетативных органов. Корень эволюционно возник несколько позднее побега и, вероятно, произошел от корнеподобных веточек-ризомоидов первых растений, вышедших на сушу. Подобные ризомоиды сохранились только у небольшой группы современных растений, относящихся к очень архаичному отделу псилотовых.
Независимо от формы тела во всех группах наземных растений очень рано (с середины девона) начала проявляться тенденция к образованию плоских боковых фотосинтезирующих органов - листьев. Подобно корням листья возникали разными путями, т.е. в разных систематических группах они имеют разное происхождение. Листья возникли двумя путями. Листья-микрофилы – появились как уплощенные выросты на теломах. Подобные листья сохранились у современных плаунов и мхов. При вхождении проводящего пучка из такого листа в стебель не образуется листовых прорывов. Листья-макрофилы возникли в результате уплощения и срастания (кладодификации) боковых веток-теломов. Такие листья у современных папоротников, хвощей и семенных растений. При вхождении листового следа в стебель образуются прорывы, как и для боковых побегов. Такую версию возникновения органов высших растений предложил в
■ наличие многоклеточных органов размножения — полового (гаметангий) и бесполого (спорангий). Мужские гаметангии высших растений называются антеридиями, женские — архегониями. Гаметангии высших растений (в отличие от низших) защищены оболочками из стерильных (бесплодных) клеток и (у отдельных групп растений) могут быть редуцированы, т.е. уменьшены и упрощены;
■ превращение зиготы в типичный многоклеточный зародыш, клетки которого первоначально не дифференцированы, но генетически детерминированы на специализацию в определенном направлении;
■ правильное чередование двух поколений — гаплоидного полового (гаметофит), развивающегося из споры, и диплоидного бесполого (спорофит), развивающегося из зиготы;
■ доминирование в жизненном цикле спорофита (у всех отделов кроме Моховидных);
■ разделение тела спорофита (у большинства отделов высших растений) на специализированные вегетативные органы — корень, стебель и листья.
Отделы:
Риниофиты (полностью исчезли)
Моховидные (25 000 видов)
Плауновидные (1 200 видов)
Хвощевидные (30 видов)
Папоротниковидные (10 000 видов)
Голосеменные (около 700 видов)
Покрытосеменные, или Цветковые (250 000 видов;)
Отличия высших растений от водорослей
-
Дифференциация растения на органы – стебель, корни, листья, репродуктивные органы -
Наличие разнообразных тканей -
Чередование полового и неполового поколение
-
Происхождение высших растений и время появления в геологической летописи. Предполагаемые предки. Приспособления высших растений к наземно-воздушной среде обитания.
Высшие растения, вероятно, произошли от каких-либо водорослей. Об этом свидетельствует то, что в геологической истории растительного мира высшим растениям предшествовали водоросли. В пользу этого предположения свидетельствуют и следующие факты: сходство наиболее древней вымершей группы высших растений - риниофитов – с водорослями, очень сходный характер их ветвления; сходство в чередовании поколений высших растений и многих водорослей; наличие жгутиков и способность к самостоятельному плаванию у
мужских половых клеток многих высших растений; сходство в строении и функции хлоропластов.
Предполагают, что высшие растения произошли скорее всего от зеленых водорослей, пресноводных или солоноватоводных. Они имели многоклеточные гаметангии, изоморфное чередование поколений в цикле развития.
Первыми наземными растениями, найденными в ископаемом состоянии, были риниофиты (риния, хорнея, хорнеофитон, спорогонитес, псилофит и др.).
После выхода на сушу высшие растения развивались в двух основных направлениях и образовали две большие эволюционные ветви – гаплоидную и диплоидную.
По имеющимся палеонтологическим данным, это произошло примерно 440 млн лет назад (силурийский период палеозойской эры). Известна точка зрения о том, что некие водоросли, жившие вблизи берега, сначала «высунули голову на воздух», затем заселили приливно-отливную зону, а потом, постепенно превращаясь в высшие растения, целиком вышли на берег. Впоследствии осуществилось постепенное завоевание ими суши.
С выходом растений на сушу у них появились ткани. В связи с тем, что воздух не может служить надежной опорой, у растений возникли механические ткани (древесные и лубяные волокна). Широкий диапазон изменения климатических факторов стал причиной формирования плотных покровных тканей (перидермы и корки). Нахождение воды и питательных веществ в почве обусловило появление проводящих тканей (ксилемы и флоэмы), формирование корня, стебля и листьев. Благодаря подвижности воздуха (ветру) у растений сформировались приспособления к опылению, распространению спор, пыльцы, плодов и семян.
-
Исторические этапы развития растительного мира на Земле. Характеристика растительного покрова в периоды палеозоя, мезозоя и кайнозоя.
Палеозой:
В первых двух периодах эры палеозоя растительный мир был представлен преимущественно водорослями. В период силура появляются первые споровые растения, а в начале делура уже имеется множество простейших растений – риниофитов. К середине этого периода растительность развивается. Появились первые плауновые, прапапоротники, членистостебельные, прогимноспермы, голосеменные. Развивается покров почвы. Карбон ознаменовал появление хвощеподобных, древовидных плацновых, папоротников и папоротникообразных, кордаитов. Флора карбона со временем образовала толстый слой каменного угля, который добывается по сей день.
Мезозой:
С окончанием палеозоя вымирают папоротники, плауны и древесные хвощи. На смену им в триасовом периоде приходят хвойные и другие голосеменные растения. В юрском периоде вымирают уже голосеменные папоротники и появляются древесные покрытосеменные растения. Всю сушу покрывает обильная растительность, появляются предшественники сосен, кипарисов, мамонтовых деревьев. В меловом периоде развились первые растения с цветами. У них был тесный контакт с насекомыми, одни без других, по сути, не существовали. Поэтому за короткое время они распространились по всем уголкам планеты.
Палеозой:
На высоких широтах начинают преобладать покрытосеменные растения и хвойные. Зона экватора была покрыта дождевыми влажными лесами (пальмы, сандал, фикусы). В глубине материковых зон были распространены саванны и редкие леса. На средних широтах росли растения тропического типа – хлебные деревья, древовидные папоротники, банановые деревья, сандал.
Заполярье было покрыто широколиственными и хвойными деревьями. В неогене начинает развиваться флора современного Средиземного моря. На севере уже почти не было вечнозеленых растений. Выделяются таежная, тундровая и лесостепная зона. На месте саванн появляются пустыни или полупустыни.
-
Отдел Rhyniophyta, время их жизни, особенности строения, филогенетическое значение.
РИНИОФИТЫ, первые наземные растения. Существовали в силуре – верхнем девоне. Примитивные сосудистые растения выс. 20–70 см, иногда до 3 м, без корней и листьев, с протостелическим (единый центральный проводящий пучок ксилемы, окружённый кольцом флоэмы) стеблем, преимущественно с дихотомическим вильчатым ветвлением и спорангиями на концах побегов, реже – вдоль побегов. Возможно, что спорофит и гаметофит существовали в виде самостоятельных растений. Росли по берегам водоёмов, располагаясь частично в воде или на насыщенных водой участках суши. Происхождение не ясно. Считаются предками других высших растений. Выход риниофитов на сушу – одно из важнейших событий в эволюции биосферы.
Риниофиты характеризовались верхушечным (терминальным) расположением спорангиев, отсутствием корней и листьев, дихотомическим, или дихоподиальным (псевдомоноподиальным), ветвлением спорофита, очень примитивным анатомическим строением и равноспоровостью. Проводящая система была слабо развита и представляла собой типичную протостелу. При развитии ксилемы первыми формировались трахеиды в центре ксилемы, т. е. ксилема была центрархная.
Филогенетическое значение: Риниофиты сам древн гр раст дала начало трем ветвям плауновидные, хвощевидные, папоротниковидные. От папоротников берут начало голосеменные, а от них цветковые. Благодаря им возникла теломная или эволюционная морфология, кот заним-ся изучением вегетативных органов высш раст.
-
Типы строения тела у высших растений. Теломный уровень строения. Особенности вегетативных и генеративных органов высших растений, в связи с выходом на сушу.
Типы органов высших растений:
■ вегетативные (корень, стебель, лист);
■ генеративные (спорангии, антеридии, архегонии, цветок).
❖ Вегетативные органы — органы, выполняющие основные функции питания и обмена веществ с внешней средой, обеспечивая фотосинтез, водоснабжение, дыхание и рост растения.
Особенности вегетативных органов. Эти органы:
■ образуют систему побегов и корневую систему;
■ не участвуют в спорообразовании и половом размножении;
■ могут выполнять функцию вегетативного размножения;
■ при изменении функций претерпевают метаморфозы (видоизменяются).
❖ Генеративные (или репродуктивные) органы — специальные органы, обеспечивающие размножение растений. Генеративные органы бесполого размножения растений:
■ у споровых растений — спорангии,
■ у семенных растений — пыльцевой мешок и семязачаток.
Генеративные органы полового размножения растений:
■ мхов и папоротников — антеридии и архегонии,
■ цветковых растений — цветки, плоды, семена.
Первые достоверные наземные растения, известные только по спорам, датируются началом силурийского периода. Из верхнесилурийских и нижнедевонских отложений по сохранившимся макроостаткам или по отпечаткам органов описаны наземные растения. Эти первые известные нам высшие растения объединены в группу риниофитов. Несмотря на анатомическую и морфологическую простоту строения, это были уже типичные наземные растения. Об этом свидетельствует наличие кутинизированной эпидермы с устьицами, развитой водопроводящей системы, состоящей из трахеид, и наличие многоклеточных спорангиев с кутинизированными спорами. Следовательно, можно предположить, что процесс освоения суши растениями начался значительно раньше - в кембрии или в ордовике. В ходе длительного эволюционного процесса возникали новые виды, постепенно формировавшие типичные наземные растения
Исторически вегетативные органы высших растений возникли несколько позднее, чем ткани. Как указывалось ранее, тканевая дифференциация таллома началась при освоении суши, как следствие перехода растений к постоянной жизни в двух средах - воздушной и почвенной. Ткани имелись уже у древнейших наземных растений риниофитов. Однако само по себе вегетативное тело риниофитов представляло более или менее разветвленную безлистную ось. Отдельные цилиндрические веточки этой оси получили название теломов. Считается, что из этих веточек-теломов возникает побег. У древнейших высших растений побег и его видоизменения выполняли все основные функции вегетативных органов. Корень эволюционно возник несколько позднее побега и, вероятно, произошел от корнеподобных веточек-ризомоидов первых растений, вышедших на сушу. Подобные ризомоиды сохранились только у небольшой группы современных растений, относящихся к очень архаичному отделу псилотовых.
Независимо от формы тела во всех группах наземных растений очень рано (с середины девона) начала проявляться тенденция к образованию плоских боковых фотосинтезирующих органов - листьев. Подобно корням листья возникали разными путями, т.е. в разных систематических группах они имеют разное происхождение. Листья возникли двумя путями. Листья-микрофилы – появились как уплощенные выросты на теломах. Подобные листья сохранились у современных плаунов и мхов. При вхождении проводящего пучка из такого листа в стебель не образуется листовых прорывов. Листья-макрофилы возникли в результате уплощения и срастания (кладодификации) боковых веток-теломов. Такие листья у современных папоротников, хвощей и семенных растений. При вхождении листового следа в стебель образуются прорывы, как и для боковых побегов. Такую версию возникновения органов высших растений предложил в