ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 22.11.2023
Просмотров: 550
Скачиваний: 3
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
1930 г. В. Циммерман. Она получила название теломной теории. Выход растений на сушу сопровождался улучшением их освещения, что активизировало процесс фотосинтеза. Это приводило к увеличению количества ассимилятов, а следовательно, к увеличению объема растений, что вызвало необходимость их морфологического
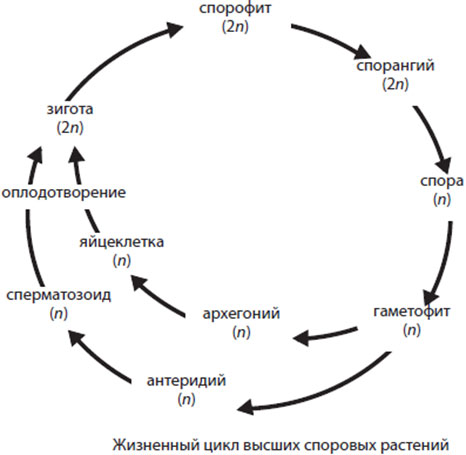
Всем высшим растением свойственно наличие двух жизненных фаз, закономерно сменяющих друг друга – гаметофита и спорофита. Вместе они составляют жизненный цикл высшего растения.
Гаметофит развивается из споры и представляет собой поколение, способное размножаться половым путем, т.е. слиянием двух гаплоидных клеток – гамет с образованием диплоидной зиготы. Поскольку гаметы у высших растений всегда образуются в результате митоза само тело гаметофита построено тоже из гаплоидных клеток.
Спорофит развивается из зиготы. Он размножается бесполым путем (спорами), и его тело образовано клетками с диплоидным набором хромосом. Спорофит не образует половые органы, и споры развиваются в особых многоклеточных структурах – спорангиях. Несмотря на то, что спорофит образован диплоидными клетками, споры гаплоидны т.к. образуются в процессе мейоза из материнских клеток – предшественников.
Органы растений, выполняющие функцию полового размножения.
У различных представителей растительного царства они сильно отличаются по уровню дифференциации и сложности строения. У некоторых водорослей и лишайников генеративные органы не дифференцированы на мужские и женские (морфологически не отличимы) и различаются только физиологически. В других случаях у низших растений генеративные органы подразделяются на антеридии (образующие мужские гаметы) и оогонии (образующие женские гаметы), как правило, одноклеточные. От них в процессе эволюции произошли многоклеточные генеративные органы высших растений - антеридии и архегонии. У высших споровых растений (мхов, папоротников, хвощей и плаунов) антеридии представляют собой небольшие овальные или шаровидные тельца, одетые снаружи бесплодными клетками, в. которых образуются подвижные сперматозоиды (мужские гаметы) (см. Мхи, Папоротниковидные, Плауновидные, Хвощевидные). Архегонии выглядят как небольшие бутылкообразные или колбообразные тельца, состоящие из брюшка и шейки. В брюшке помещается неподвижная яйцеклетка - женская гамета. В процессе эволюции высших растений архегонии и антеридии претерпевают упрощение (редукцию). У наиболее высокоорганизованных покрытосеменных растений, а также у некоторых голосеменных растений архегонии отсутствуют. У сосновых от архегония сохраняется яйцеклетка и несколько боковых клеток, а мужской гаметофит редуцирован до трех клеток и антеридии, и как таковой не образуется. У покрытосеменных (цветковых) растений, в связи с особенностями их развития, возникают высокоспециализированные генеративные органы - мужские гаметофиты (пыльцевые зерна), состоящие из двух клеток (генеративной и вегетативной), и сильно редуцированные женские гаметофиты - зародышевые мешки. Понятие Генеративные органы, у покрытосеменных растений часто распространяют на цветки и плоды.
Жизненный цикл высших растений состоит из двух фаз (или поколений) - бесполой, или спорофазы (спорофита), и половой, или гаметофазы (гаметофита). У всех высших растений, кроме мохообразных, в жизненном цикле преобладает спорофит, гаметофит развит слабее и относительно недолговечен. В жизненном цикле мохообразных преобладает гаметофит.
В эволюции остальных высших растений происходит постепенная редукция (уменьшение и упрощение) гаметофита и преобладание в жизненном цикле спорофита.
Спорофиты большинства отделов высших растений (плауновидных, хвощевидных, папоротниковидных, голосеменных и покрытосеменных) представляют собой крупные многоклеточные организмы со сложным анатомическим строением и расчленением тела на органы - стебли, листья, корни (настоящие или придаточные). Гаметофит у плауновидных, хвощевидных и папоротниковидных представлен заростком - маленьким (от нескольких мм до 3 см), не расчлененным на органы растеньицем, живущим несколько недель (у плаунов — несколько лет) независимо от спорофита
У голо- и покрытосеменных растений гаметофит полностью утратил свою самостоятельность, и все его развитие протекает на спорофите внутри макроспорангия (или нуцеллуса семяпочки). Споры (n)->геметофит (n)->митоз ->мужская и женская гамета(n)->оплодотворение->зигота(2n)->спорофит (2n)->редукционное деление->споры
Гаметофит-половое поколение
Спорофит-бесполое поколение
+Чередование поколений может быть изоморфным(поколения в жизненном цикле представлены одинаково) и гетероморфным( одно из поколений доминирует).
Эволюция жизненного цикла высших растений шла в двух противоположных направлениях. У моховидных она была направлена в сторону возрастания самостоятельности гаметофита и его постепенного морфологического расчленения, потери самостоятельности спорофита и его морфологического упрощения. Самостоятельной, вполне автотрофной фазой жизненного цикла моховидных стал гаметофит, а спорофит низведен до степени органа гаметофита. У всех остальных высших растений самостоятельной фазой жизненного цикла стал спорофит, а гаметофит у них в течение эволюции постепенно уменьшался и упрощался. Максимальная редукция гаметофита связана с разделением полов. Миниатюризация и упрощение однополых гаметофитов происходили весьма ускоренными темпами. Гаметофиты очень быстро теряли хлорофилл, и развитие все чаще осуществлялось за счет питательных веществ, накопленных спорофитом.
Разноспо́ровые расте́ния (также гетероспо́ровые расте́ния) — растения, образующие два вида спор: мужские микроспоры (меньшего размера) и женские мегаспоры (крупнее).
Гетероспоровыми является часть папоротников, плауновидные, членистостебельные, а также все голосеменные и покрытосеменные растения.
Противоположностью разноспоровости (гетероспории) является равноспоровость (изоспория), когда споры имеют одинаковый размер.
К равноспоровым растениям относятся моховидные, псилотовидные, группа плауновидных, хвощевидные и группа папоротниковидных.
Биологическое и эволюционное преимущество разноспоровости перед равноспоровостью связано с увеличением разнообарзия генетического материала, который участвует в отборе, поскольку при равноспоровости и мужские и женские гаметы образуются на одном гаметофите, а при разноспоровости из разных спор развиваются самостоятельные гаметофиты – мужские заростки и женские заростки.
Мохообразные - это уникальная группа высших растений, которые характеризуются преобладанием в цикле воспроизведения гаплоидного гаметофита над диплоидным спорофитом. Это объясняется прежде всего тем, что гаметофит мохообразных составляет основное тело растения. Для осуществления полового процесса, который происходит у них с помощью сперматозоидов, необходима влажная среда. Следовательно, мохообразные вынуждены находиться в приземных, наиболее влажных слоях атмосферы, что и обусловливает их малые размеры. Кроме того, поскольку гаплоидный гаметофит обладает меньшим генетическим потенциалом по сравнению с диплоидным спорофитом, ему присущ менее активный процесс обмена веществ. Обладая небольшими размерами и будучи связанными с влажными местообитаниями, мохообразные с момента своего возникновения оказались под пологом более крупных растений, т.е. в условиях пониженного освещения. Поэтому смена эпох, изменения в растительном покрове земли не привнесли значительных изменений в условия жизни мохообразных. Это способствовало сохранению у них множества изначальных, т.е. примитивных, признаков.
К числу примитивных признаков мохообразных относится способность при неблагоприятных условиях переходить в состояние анабиоза, что отражает общую низкую активность обмена веществ. В состоянии анабиоза, при котором практически прекращаются все обменные процессы, растения могут длительное время (иногда десятилетиями) переносить экстремальные условия - дефицит влаги, высокие (до 70, 120 °С) или отрицательные температуры. При этом мохообразные не активно противостоят факторам среды, а пассивно «уходят» от неблагоприятных условий.
Многие мохообразные относятся к числу олиготрофов, т.е. растений, способных произрастать на почвах, бедных минеральными веществами. Это свойство, по-видимому, также связано с их пониженным обменом веществ и незначительными размерами, но оно позволяет мохообразным осваивать наиболее бедные места обитания.
В основу классификации мохообразных бриологи кладут разные признаки, но во многом она определяется их взглядами на происхождение этой группы (проблема происхождения моховидных решается также неоднозначно). Согласно наиболее распространенной классификации, мохообразные подразделяются на три класса - печеночники (или маршанциевые), антоцеро-товые и листостебельные мхи (или просто мхи). В основу этой классификации положено морфологическое строение тела гаметофита, особенности строения ризоидов, строение и характер раскрывания коробочек, а также географическое распространение. Однако представители класса антоцеротовых характеризуются настолько отличающимся строением коробочки, что некоторые биологи придают им статус самостоятельного отдела.
Класс мхи включает более 14,5 тыс. видов и объединяет 3 подкласса - сфагновые, зеленые и андреевы мхи.
Подкласс сфагновые, белые, или торфяные мхи (Sphagnidae). Этот подкласс представлен одним порядком Sphagnales, одним семейством Sphagnaceae с одним родом сфагнум (Sphagnum), объединяющим свыше 300 видов, которые морфологически довольно трудно различаются. В России произрастает 42 вида. Все представители беловато-зеленого цвета, не имеют ризоидов. От стебля растения отходят боковые веточки, на верхушке собранные в головку.
Побеги сфагнума усажены мелкими листьями. Листья однослойные, состоят из двух типов клеток - хлорофиллоносных и мертвых водоносных (гиалиновых). За счет наличия в листе мертвых гиалиновых клеток, способных удерживать воду, они обладают гигроскопическим
свойством. Фотосинтез протекает в живых ассимиляционных клетках, содержащих хлоропласты (рис.3, Б). При высыхании мертвые клетки заполняются воздухом, и мох становится беловатым, за счет чего и получил название белый мох.
Подкласс андреевые мхи (andreaeidae). Андреевые мхи - это небольшая группа мхов, представленная одним семейством Andreaeaceae с единственным родом андреа (Andreaea), или андреев мох, объединяющим 120 видов.
Подкласс зеленые мхи (Bryidae). К числу особенностей этого подкласса относятся: 1) наличие хорошо развитой нитчатой протонемы, на которой формируются побеги; 2) наличие в листе средней жилки, даже если листовая пластинка однослойная; 3) сложное строение спорогония, состоящего из коробочки, ножки и гаустории; 4) наличие в коробочке перистома и колонки, доходящей до вершины.
Характерным представителем подкласса является мох кукушкин лен обыкновенный (Polytrchum commune). Кукушкин лен - один из наиболее высокорослых мхов, его стебель достигает 50 см в высоту. Произрастает на сырой почве в лесах и болотах, образуя крупные подушкообразные дернины.
- Класс Hepaticopsida:
Класс Печеночные мхи.
- Особенности строения:
Всегда билатеральная симметрия, дорзовентральное строение.
Талломы могут быть пластинчатыми (слоевищными). У большинства видов класса талломы листостебельные. На нижней стороне располагаются ризоиды и однослойные выросты таллома – амфигастрии. Ризоиды только одноклеточные, разнообразного строения (простые, язычковые, иногда разветвленные). Листья разнообразной формы, часто разделены на лопасти, обычно без жилки и однослойные. Листорасположение двухстороннее, очередное и реже супротивное. В клетках таллома и листьев имеются характерные масляные тельца от одного, нескольких, или до полусотни.
У печеночных мхов широко распространено вегетативное размножение как частями тела, так и специализированными органами: выводковыми почками (геммы), выводковыми тельцами, придаточными побегами, ломкими листьями, клубеньками и другими приспособлениями.
Гаметангии могут располагаться на особых выростах таллома (подставках), либо на самих талломах.
Диплоидный спорофит печеночных мхов состоит обычно из коробочки, ножки и гаустории. Спорогон без хлоропластов, колонки и устьиц. Коробочка функционально является спорангием. Важную функцию разрыхления спор выполняют элатеры (пружинки), формирующиеся у большинства видов.
-
Характеристика цикла воспроизведения высших растений, его варианты, главные направления эволюции.
Всем высшим растением свойственно наличие двух жизненных фаз, закономерно сменяющих друг друга – гаметофита и спорофита. Вместе они составляют жизненный цикл высшего растения.
Гаметофит развивается из споры и представляет собой поколение, способное размножаться половым путем, т.е. слиянием двух гаплоидных клеток – гамет с образованием диплоидной зиготы. Поскольку гаметы у высших растений всегда образуются в результате митоза само тело гаметофита построено тоже из гаплоидных клеток.
Спорофит развивается из зиготы. Он размножается бесполым путем (спорами), и его тело образовано клетками с диплоидным набором хромосом. Спорофит не образует половые органы, и споры развиваются в особых многоклеточных структурах – спорангиях. Несмотря на то, что спорофит образован диплоидными клетками, споры гаплоидны т.к. образуются в процессе мейоза из материнских клеток – предшественников.
Органы растений, выполняющие функцию полового размножения.
У различных представителей растительного царства они сильно отличаются по уровню дифференциации и сложности строения. У некоторых водорослей и лишайников генеративные органы не дифференцированы на мужские и женские (морфологически не отличимы) и различаются только физиологически. В других случаях у низших растений генеративные органы подразделяются на антеридии (образующие мужские гаметы) и оогонии (образующие женские гаметы), как правило, одноклеточные. От них в процессе эволюции произошли многоклеточные генеративные органы высших растений - антеридии и архегонии. У высших споровых растений (мхов, папоротников, хвощей и плаунов) антеридии представляют собой небольшие овальные или шаровидные тельца, одетые снаружи бесплодными клетками, в. которых образуются подвижные сперматозоиды (мужские гаметы) (см. Мхи, Папоротниковидные, Плауновидные, Хвощевидные). Архегонии выглядят как небольшие бутылкообразные или колбообразные тельца, состоящие из брюшка и шейки. В брюшке помещается неподвижная яйцеклетка - женская гамета. В процессе эволюции высших растений архегонии и антеридии претерпевают упрощение (редукцию). У наиболее высокоорганизованных покрытосеменных растений, а также у некоторых голосеменных растений архегонии отсутствуют. У сосновых от архегония сохраняется яйцеклетка и несколько боковых клеток, а мужской гаметофит редуцирован до трех клеток и антеридии, и как таковой не образуется. У покрытосеменных (цветковых) растений, в связи с особенностями их развития, возникают высокоспециализированные генеративные органы - мужские гаметофиты (пыльцевые зерна), состоящие из двух клеток (генеративной и вегетативной), и сильно редуцированные женские гаметофиты - зародышевые мешки. Понятие Генеративные органы, у покрытосеменных растений часто распространяют на цветки и плоды.
Жизненный цикл высших растений состоит из двух фаз (или поколений) - бесполой, или спорофазы (спорофита), и половой, или гаметофазы (гаметофита). У всех высших растений, кроме мохообразных, в жизненном цикле преобладает спорофит, гаметофит развит слабее и относительно недолговечен. В жизненном цикле мохообразных преобладает гаметофит.
В эволюции остальных высших растений происходит постепенная редукция (уменьшение и упрощение) гаметофита и преобладание в жизненном цикле спорофита.
Спорофиты большинства отделов высших растений (плауновидных, хвощевидных, папоротниковидных, голосеменных и покрытосеменных) представляют собой крупные многоклеточные организмы со сложным анатомическим строением и расчленением тела на органы - стебли, листья, корни (настоящие или придаточные). Гаметофит у плауновидных, хвощевидных и папоротниковидных представлен заростком - маленьким (от нескольких мм до 3 см), не расчлененным на органы растеньицем, живущим несколько недель (у плаунов — несколько лет) независимо от спорофита
У голо- и покрытосеменных растений гаметофит полностью утратил свою самостоятельность, и все его развитие протекает на спорофите внутри макроспорангия (или нуцеллуса семяпочки). Споры (n)->геметофит (n)->митоз ->мужская и женская гамета(n)->оплодотворение->зигота(2n)->спорофит (2n)->редукционное деление->споры
Гаметофит-половое поколение
Спорофит-бесполое поколение
+Чередование поколений может быть изоморфным(поколения в жизненном цикле представлены одинаково) и гетероморфным( одно из поколений доминирует).
Эволюция жизненного цикла высших растений шла в двух противоположных направлениях. У моховидных она была направлена в сторону возрастания самостоятельности гаметофита и его постепенного морфологического расчленения, потери самостоятельности спорофита и его морфологического упрощения. Самостоятельной, вполне автотрофной фазой жизненного цикла моховидных стал гаметофит, а спорофит низведен до степени органа гаметофита. У всех остальных высших растений самостоятельной фазой жизненного цикла стал спорофит, а гаметофит у них в течение эволюции постепенно уменьшался и упрощался. Максимальная редукция гаметофита связана с разделением полов. Миниатюризация и упрощение однополых гаметофитов происходили весьма ускоренными темпами. Гаметофиты очень быстро теряли хлорофилл, и развитие все чаще осуществлялось за счет питательных веществ, накопленных спорофитом.
-
Равно- и разноспоровые растения. Биологическое и эволюционное значение перехода от равноспоровости к разноспоровости у высших растений.
Разноспо́ровые расте́ния (также гетероспо́ровые расте́ния) — растения, образующие два вида спор: мужские микроспоры (меньшего размера) и женские мегаспоры (крупнее).
Гетероспоровыми является часть папоротников, плауновидные, членистостебельные, а также все голосеменные и покрытосеменные растения.
Противоположностью разноспоровости (гетероспории) является равноспоровость (изоспория), когда споры имеют одинаковый размер.
К равноспоровым растениям относятся моховидные, псилотовидные, группа плауновидных, хвощевидные и группа папоротниковидных.
Биологическое и эволюционное преимущество разноспоровости перед равноспоровостью связано с увеличением разнообарзия генетического материала, который участвует в отборе, поскольку при равноспоровости и мужские и женские гаметы образуются на одном гаметофите, а при разноспоровости из разных спор развиваются самостоятельные гаметофиты – мужские заростки и женские заростки.
-
Общая характеристика и принципы классификации мохообразных. Различные взгляды на их таксономический статус.
Мохообразные - это уникальная группа высших растений, которые характеризуются преобладанием в цикле воспроизведения гаплоидного гаметофита над диплоидным спорофитом. Это объясняется прежде всего тем, что гаметофит мохообразных составляет основное тело растения. Для осуществления полового процесса, который происходит у них с помощью сперматозоидов, необходима влажная среда. Следовательно, мохообразные вынуждены находиться в приземных, наиболее влажных слоях атмосферы, что и обусловливает их малые размеры. Кроме того, поскольку гаплоидный гаметофит обладает меньшим генетическим потенциалом по сравнению с диплоидным спорофитом, ему присущ менее активный процесс обмена веществ. Обладая небольшими размерами и будучи связанными с влажными местообитаниями, мохообразные с момента своего возникновения оказались под пологом более крупных растений, т.е. в условиях пониженного освещения. Поэтому смена эпох, изменения в растительном покрове земли не привнесли значительных изменений в условия жизни мохообразных. Это способствовало сохранению у них множества изначальных, т.е. примитивных, признаков.
К числу примитивных признаков мохообразных относится способность при неблагоприятных условиях переходить в состояние анабиоза, что отражает общую низкую активность обмена веществ. В состоянии анабиоза, при котором практически прекращаются все обменные процессы, растения могут длительное время (иногда десятилетиями) переносить экстремальные условия - дефицит влаги, высокие (до 70, 120 °С) или отрицательные температуры. При этом мохообразные не активно противостоят факторам среды, а пассивно «уходят» от неблагоприятных условий.
Многие мохообразные относятся к числу олиготрофов, т.е. растений, способных произрастать на почвах, бедных минеральными веществами. Это свойство, по-видимому, также связано с их пониженным обменом веществ и незначительными размерами, но оно позволяет мохообразным осваивать наиболее бедные места обитания.
В основу классификации мохообразных бриологи кладут разные признаки, но во многом она определяется их взглядами на происхождение этой группы (проблема происхождения моховидных решается также неоднозначно). Согласно наиболее распространенной классификации, мохообразные подразделяются на три класса - печеночники (или маршанциевые), антоцеро-товые и листостебельные мхи (или просто мхи). В основу этой классификации положено морфологическое строение тела гаметофита, особенности строения ризоидов, строение и характер раскрывания коробочек, а также географическое распространение. Однако представители класса антоцеротовых характеризуются настолько отличающимся строением коробочки, что некоторые биологи придают им статус самостоятельного отдела.
Класс мхи включает более 14,5 тыс. видов и объединяет 3 подкласса - сфагновые, зеленые и андреевы мхи.
Подкласс сфагновые, белые, или торфяные мхи (Sphagnidae). Этот подкласс представлен одним порядком Sphagnales, одним семейством Sphagnaceae с одним родом сфагнум (Sphagnum), объединяющим свыше 300 видов, которые морфологически довольно трудно различаются. В России произрастает 42 вида. Все представители беловато-зеленого цвета, не имеют ризоидов. От стебля растения отходят боковые веточки, на верхушке собранные в головку.
Побеги сфагнума усажены мелкими листьями. Листья однослойные, состоят из двух типов клеток - хлорофиллоносных и мертвых водоносных (гиалиновых). За счет наличия в листе мертвых гиалиновых клеток, способных удерживать воду, они обладают гигроскопическим
свойством. Фотосинтез протекает в живых ассимиляционных клетках, содержащих хлоропласты (рис.3, Б). При высыхании мертвые клетки заполняются воздухом, и мох становится беловатым, за счет чего и получил название белый мох.
Подкласс андреевые мхи (andreaeidae). Андреевые мхи - это небольшая группа мхов, представленная одним семейством Andreaeaceae с единственным родом андреа (Andreaea), или андреев мох, объединяющим 120 видов.
Подкласс зеленые мхи (Bryidae). К числу особенностей этого подкласса относятся: 1) наличие хорошо развитой нитчатой протонемы, на которой формируются побеги; 2) наличие в листе средней жилки, даже если листовая пластинка однослойная; 3) сложное строение спорогония, состоящего из коробочки, ножки и гаустории; 4) наличие в коробочке перистома и колонки, доходящей до вершины.
Характерным представителем подкласса является мох кукушкин лен обыкновенный (Polytrchum commune). Кукушкин лен - один из наиболее высокорослых мхов, его стебель достигает 50 см в высоту. Произрастает на сырой почве в лесах и болотах, образуя крупные подушкообразные дернины.
-
Класс Hepaticopsida, особенности строения, размножения, цикла воспроизведения. Важнейшие представители.
- Класс Hepaticopsida:
Класс Печеночные мхи.
- Особенности строения:
Всегда билатеральная симметрия, дорзовентральное строение.
Талломы могут быть пластинчатыми (слоевищными). У большинства видов класса талломы листостебельные. На нижней стороне располагаются ризоиды и однослойные выросты таллома – амфигастрии. Ризоиды только одноклеточные, разнообразного строения (простые, язычковые, иногда разветвленные). Листья разнообразной формы, часто разделены на лопасти, обычно без жилки и однослойные. Листорасположение двухстороннее, очередное и реже супротивное. В клетках таллома и листьев имеются характерные масляные тельца от одного, нескольких, или до полусотни.
У печеночных мхов широко распространено вегетативное размножение как частями тела, так и специализированными органами: выводковыми почками (геммы), выводковыми тельцами, придаточными побегами, ломкими листьями, клубеньками и другими приспособлениями.
Гаметангии могут располагаться на особых выростах таллома (подставках), либо на самих талломах.
Диплоидный спорофит печеночных мхов состоит обычно из коробочки, ножки и гаустории. Спорогон без хлоропластов, колонки и устьиц. Коробочка функционально является спорангием. Важную функцию разрыхления спор выполняют элатеры (пружинки), формирующиеся у большинства видов.