ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 04.12.2023
Просмотров: 128
Скачиваний: 3
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
Глава 3. Метаболизм нуклеотидов
Простейщие представители – мононуклеотиды – включают в молекулу фосфорную кислоту, пентозу и азотсодержащий компонент (обычно пуриновое или пиримидиновое основание) (3.1., приложение, табл. 4.5). Основная масса мононуклеотидов служит макроэргами, энергия которых используется для различных энергоемких процессов, в том числе для синтеза ди- , полинуклеотидов. Самые сложные полимеры – полинуклеотиды – в зависимости от особенностей строения и функций делят на РНК и ДНК. Среди РНК выделяют транспортную, ответственную за доставку активированной аминокислоты к рибосоме, а также за узнавание места расположения в аминокислотной последовательности, рибосомальную, составляющую основу рибосом и обеспечивающую образование пептидной связи, информационную – указывающую на порядок аминокислот в полипептидной цепи. По локализации (в ядре или митохондрии) делят ДНК, которые отличаются по массе, и по строению . Все полинуклеотиды обладают высокой плотностью, кислотностью, способны к денатурации и другим свойствам. По происхождению даже простейшие представители – мононуклеотиды – имеют характер. Гидролиз же пищевых полинуклеотидов начинается в желудке, заканчивается в тонком кишечнике, осуществляется с помощью специфических гидролаз: РНК-аз, ДНК-аз, щелочной фосфатазы. Пуриновые нуклеозиды преобразуются в энтероцитах или в печени в мочевую кислоту – конечный продукт катаболизма, её соли выводятся с мочой. Конечными продуктами распада пиримидиновых нуклеозидов являются метаболиты ЦТК, а также СО2, NН3, которые позднее включаются в мочевину.
Для синтеза мононуклеотидов, точнее для их пуринов и пиримидинов используются атомы или группировки атомов аминокислот, а также СО2 (3.4). Кольца пиримидинов включают аспартат, амидную группу глутамина и СО2. Полученные структуры взаимодействуют с ФРПФ с получением соответствующего мононуклеотида.
Для образования пуринов необходимы глутамин, аспартат, глицин, СО2, одноуглеродные фрагменты, переносимые ТГФК, их атомы постепенно образуют конденсированные гетероциклы на фундаменте фосфорибозы.
Полученные нуклеозидмонофосфаты дважды фосфорилируются, преобразуясь в макроэрги; рибонуклеозиддифосфаты окисляются в дезоксипроизводные, необходимые для синтеза ДНК. Из всех патологических состояний обмена нуклеотидов наиболее распространена подагра, редко встречаются наследственные болезни (3.5, «Патохимия нас. заболеваний».
Классификация и номенклатура нуклеотидов
В отличие от углеводов, липидов, белков, у которых простейшие представители их классов не способны к гидролизу, их аналогов среди изучаемых соединений – мононуклеотидов – характерна подобная реакция, т. к. они представляют сложные эфиры фосфорной кислоты и нуклеотидов. Судя по окончанию, последние являются гликозидами, где роль аглюкона выполняет какое – либо азотистое гетероциклическое основание (чаще пурин или пиримидин) (Приложение табл. 12), а в качестве углеводного компонента используется обычно пентоза (рибоза, дезоксирибоза, изредка восстановленная форма – рибитол). Таким образом, мононуклеотид может гидролизоваться по следующим связям: сложноэфирной и N–гликозидной:
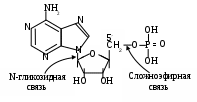
Номенклатура наиболее часто встречающихся мононуклеидов и включенных в них оснований представлена в таблицах 12, 13 (приложение). Обращают на себя разные изменения окончаний пуринов и пиримидинов при их взаимодействии с моносахаридом – продукты первых приобретают окончание – озин (аденозин, гуанозин), вторых – идин (цитидин, уридин, тимидин). Чтобы указать природу пентозы, перед основой добавляют приставку дезокси, если называемое соединение включает дезоксирибозу. В мононуклеотидах обычно фосфат связан с пятым углеродным атомом пентозы. Два звена мононуклеотида: азотистое основание и углевод – имеют несколько атомов углерода, поэтому для их опознания приняты следующие обозначения: атомы первого нумеруют арабскими цифрами, для атомов пентозы добавляют «׳» - знак (штрих) (у пятого атома штрих (см. выше)).
Особенности строения, биологическая роль нуклеиновых соединений
Функции мононуклеотидов
Большая часть нуклеотидмонофосфатов дважды фосфорилируется, используя, в основном макроэрг. – АТФ:
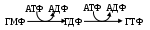
Образующиеся нуклеозидтрифосфаты несут количество энергии, достаточное для протекания самых сложных энергоемких процессов, например, синтеза различных соединений, т. е. являются макроэргами. Во – вторых, они способны полимеризоваться в нуклеиновые кислоты или динуклеотиды. Разница в путях образования следующая: наиболее реакционно способен фосфат монопроизводного. Если реагируют между собой фосфаты обоих мононуклеотидов, то получившийся динуклеотид оказывается стабильным и используется клетками в качестве кофермента (НАД+, НАД+Ф, ФАД). Если же фосфат, стоящий 5׳ - углеродного атома пентозы одного мононуклеотида, реагирует с 3׳ - углеродным атомом пентозы другого мононуклеотида, то фосфат последнего способен вступить в такую же реакцию с третьим аналогом. Так и происходит полимеризация отдельных звеньев в мононуклеотиды.
Кроме того, в плазмолеммах имеется особая ферментная – аденилатциклазная – система, которая под влиянием различных биологически активных веществ (гормонов) преобразует АТФ в ц-АМФ – служащий secondmessenger (вторичным посредником) – переводящим сигнал гормона внутрь клетки. А еще описаны мононуклеотиды, выполняющие коферментные функции (ФМН – флавинмононуклеотид–коэнзим ДГ).
Как частный случай можно представить продукт взаимодействия АТФ с аминокислотами – аминоациладенилаты (метаболиты в активации этих соединений в трансляции), S-аденозилметионин (SAM: активная форма метионина в реакциях метилирования – см. выше).
Значение динуклеотидов
Как отмечено выше, динуклеотиды обычно выполняют роль коферментов. Большая часть входит в состав дегидрогеназ (НАД+, НАД+Ф, ФАД). Еще один, единственный их представитель – НS-KoA – облигатный участник реакций ацилирования: его включение в состав любой органической кислоты (aцил
SKoA) активирует ее, увеличивает растворимость (метаболиты ЦТК, β-окисления ВЖК, гликолиза, синтеза гема и т. д.)
Полинуклеотиды
Этим термином обозначаются полимеры, мономерами которых служат мононуклеотиды. В зависимости от входящих в них пентоз их делят на рибонуклеиновые (РНК) и дезоксирибонуклеиновые (ДНК). Их нити, как и в белках, не ветвятся (в отличие от гликогена), но, как и все подобные соединения, компактно укладываются в пространстве, т. е. имеют сложную организацию (приложения, схема 2).
Первичная структура – определенная последовательность мононуклеотидов, связанных между собой фосфодиэфирной связью. Этот уровень строения полинуклеотидов отличает их друг от друга: 1) мононуклеотиды РНК включают урацил, цитозин, аденин, гуанин, а ДНК – вместо первого пиримидинового основания имеют его метилированное производное – тимин; 2) как уже описано выше, мононуклеотиды РНК и ДНК содержат разные пентозы; 3) иногда, кроме известных пуринов и пиримидинов, регистрируются их модифицированные аналоги (метилгуанин, метиладенин, дигидроурацил, метилцитозин, и т. д.) – названные минорными основаниями; разные полинуклеотиды отличаются их содержанием; 4) РНК обычно представляют одну цепь, а ДНК эукариот – это двойная нить; 5) молекулярная масса любых РНК меньше, чем ДНК; 6) содержание в клетках ДНК – величина постоянная, а РНК – лабильная; 7) отличаются нуклеиновые кислоты функциями и локализацией.
Виды РНК
Среди РНК различают: информационную, рибосомальную, транспортную, регуляторную, гетерогенно ядерную и др. На долю транспортной РНК (тРНК) приходится 10-15% всех РНК клетки, молекулярная масса не превышает 30 тыс. Д, ее нить состоит из 70-95 мононуклеотидов, причем в ее молекулу включено до 8-19% минорных оснований (дигидрацил, инозин, тиоурацил и т. д.). Необычность их структуры мешает нуклеазам разрушать тРНК, что делает их долгожителями. Оба конца нити тРНК у всех их представителей имеют одинаковое строение: в начале цепи расположен ГМФ, а заканчивается следующим тринуклеотидом: ЦЦА. Вторичная структура представлена в форме клеверного листа, образованной в виде шпилькообразных структур, за счет комплементарности (схема). Сближенные концы цепи тРНК называют акцепторным участком, сюда крепится транспортируемая аминокислота, противоположная петля включает антикодон – триплет, с помощью которого узнается соответствующее (комплементарное) звено на иРНК. Исходя из того, что смысловых кодонов 61, следовательно, и разных тРНК в клетке не менее 60. Еще две петли служат для взаимодействия с рибосомой и аминоацил-тРНК-синтетазой. Цепи различных тРНК укладываются в компактные третичные структуры; за счет стекинг - взаимодействий петли сближаются, принимая форму, напоминающую латинскую букву «L». Митохондриальные тРНК несколько короче (до 65 нуклеотидов), петли также меньше, пространственная структура не установлена.
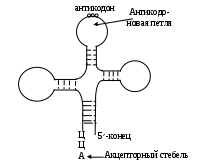
Схема 3.2.3.1.1. Универсальная укладка полинуклеотидной цепи тРНК.
Функции т-РНК: а) акцепторная – способность ковалентно связываться с аминокислотой, превращаясь в аминоацил
тРНК; б) адапторная – способность узнавать кодом иРНК, соответствующий транспортируемой аминокислоте, и обеспечивать ее включение на законное место в растущей цепи белка. При этом образование пептидной связи обеспечивается за счет макроэргической связи аминоацил
т-РНК .
Глава 3. Метаболизм нуклеотидов
Простейщие представители – мононуклеотиды – включают в молекулу фосфорную кислоту, пентозу и азотсодержащий компонент (обычно пуриновое или пиримидиновое основание) (3.1., приложение, табл. 4.5). Основная масса мононуклеотидов служит макроэргами, энергия которых используется для различных энергоемких процессов, в том числе для синтеза ди- , полинуклеотидов. Самые сложные полимеры – полинуклеотиды – в зависимости от особенностей строения и функций делят на РНК и ДНК. Среди РНК выделяют транспортную, ответственную за доставку активированной аминокислоты к рибосоме, а также за узнавание места расположения в аминокислотной последовательности, рибосомальную, составляющую основу рибосом и обеспечивающую образование пептидной связи, информационную – указывающую на порядок аминокислот в полипептидной цепи. По локализации (в ядре или митохондрии) делят ДНК, которые отличаются по массе, и по строению . Все полинуклеотиды обладают высокой плотностью, кислотностью, способны к денатурации и другим свойствам. По происхождению даже простейшие представители – мононуклеотиды – имеют характер. Гидролиз же пищевых полинуклеотидов начинается в желудке, заканчивается в тонком кишечнике, осуществляется с помощью специфических гидролаз: РНК-аз, ДНК-аз, щелочной фосфатазы. Пуриновые нуклеозиды преобразуются в энтероцитах или в печени в мочевую кислоту – конечный продукт катаболизма, её соли выводятся с мочой. Конечными продуктами распада пиримидиновых нуклеозидов являются метаболиты ЦТК, а также СО2, NН3, которые позднее включаются в мочевину.
Для синтеза мононуклеотидов, точнее для их пуринов и пиримидинов используются атомы или группировки атомов аминокислот, а также СО2 (3.4). Кольца пиримидинов включают аспартат, амидную группу глутамина и СО2. Полученные структуры взаимодействуют с ФРПФ с получением соответствующего мононуклеотида.
Для образования пуринов необходимы глутамин, аспартат, глицин, СО2, одноуглеродные фрагменты, переносимые ТГФК, их атомы постепенно образуют конденсированные гетероциклы на фундаменте фосфорибозы.
Полученные нуклеозидмонофосфаты дважды фосфорилируются, преобразуясь в макроэрги; рибонуклеозиддифосфаты окисляются в дезоксипроизводные, необходимые для синтеза ДНК. Из всех патологических состояний обмена нуклеотидов наиболее распространена подагра, редко встречаются наследственные болезни (3.5, «Патохимия нас. заболеваний».
Классификация и номенклатура нуклеотидов
В отличие от углеводов, липидов, белков, у которых простейшие представители их классов не способны к гидролизу, их аналогов среди изучаемых соединений – мононуклеотидов – характерна подобная реакция, т. к. они представляют сложные эфиры фосфорной кислоты и нуклеотидов. Судя по окончанию, последние являются гликозидами, где роль аглюкона выполняет какое – либо азотистое гетероциклическое основание (чаще пурин или пиримидин) (Приложение табл. 12), а в качестве углеводного компонента используется обычно пентоза (рибоза, дезоксирибоза, изредка восстановленная форма – рибитол). Таким образом, мононуклеотид может гидролизоваться по следующим связям: сложноэфирной и N–гликозидной:
Номенклатура наиболее часто встречающихся мононуклеидов и включенных в них оснований представлена в таблицах 12, 13 (приложение). Обращают на себя разные изменения окончаний пуринов и пиримидинов при их взаимодействии с моносахаридом – продукты первых приобретают окончание – озин (аденозин, гуанозин), вторых – идин (цитидин, уридин, тимидин). Чтобы указать природу пентозы, перед основой добавляют приставку дезокси, если называемое соединение включает дезоксирибозу. В мононуклеотидах обычно фосфат связан с пятым углеродным атомом пентозы. Два звена мононуклеотида: азотистое основание и углевод – имеют несколько атомов углерода, поэтому для их опознания приняты следующие обозначения: атомы первого нумеруют арабскими цифрами, для атомов пентозы добавляют «׳» - знак (штрих) (у пятого атома штрих (см. выше)).
Особенности строения, биологическая роль нуклеиновых соединений
Функции мононуклеотидов
Большая часть нуклеотидмонофосфатов дважды фосфорилируется, используя, в основном макроэрг. – АТФ:
Образующиеся нуклеозидтрифосфаты несут количество энергии, достаточное для протекания самых сложных энергоемких процессов, например, синтеза различных соединений, т. е. являются макроэргами. Во – вторых, они способны полимеризоваться в нуклеиновые кислоты или динуклеотиды. Разница в путях образования следующая: наиболее реакционно способен фосфат монопроизводного. Если реагируют между собой фосфаты обоих мононуклеотидов, то получившийся динуклеотид оказывается стабильным и используется клетками в качестве кофермента (НАД+, НАД+Ф, ФАД). Если же фосфат, стоящий 5׳ - углеродного атома пентозы одного мононуклеотида, реагирует с 3׳ - углеродным атомом пентозы другого мононуклеотида, то фосфат последнего способен вступить в такую же реакцию с третьим аналогом. Так и происходит полимеризация отдельных звеньев в мононуклеотиды.
Кроме того, в плазмолеммах имеется особая ферментная – аденилатциклазная – система, которая под влиянием различных биологически активных веществ (гормонов) преобразует АТФ в ц-АМФ – служащий secondmessenger (вторичным посредником) – переводящим сигнал гормона внутрь клетки. А еще описаны мононуклеотиды, выполняющие коферментные функции (ФМН – флавинмононуклеотид–коэнзим ДГ).
Как частный случай можно представить продукт взаимодействия АТФ с аминокислотами – аминоациладенилаты (метаболиты в активации этих соединений в трансляции), S-аденозилметионин (SAM: активная форма метионина в реакциях метилирования – см. выше).
Значение динуклеотидов
Как отмечено выше, динуклеотиды обычно выполняют роль коферментов. Большая часть входит в состав дегидрогеназ (НАД+, НАД+Ф, ФАД). Еще один, единственный их представитель – НS-KoA – облигатный участник реакций ацилирования: его включение в состав любой органической кислоты (aцил
SKoA) активирует ее, увеличивает растворимость (метаболиты ЦТК, β-окисления ВЖК, гликолиза, синтеза гема и т. д.)Полинуклеотиды
Этим термином обозначаются полимеры, мономерами которых служат мононуклеотиды. В зависимости от входящих в них пентоз их делят на рибонуклеиновые (РНК) и дезоксирибонуклеиновые (ДНК). Их нити, как и в белках, не ветвятся (в отличие от гликогена), но, как и все подобные соединения, компактно укладываются в пространстве, т. е. имеют сложную организацию (приложения, схема 2).
Первичная структура – определенная последовательность мононуклеотидов, связанных между собой фосфодиэфирной связью. Этот уровень строения полинуклеотидов отличает их друг от друга: 1) мононуклеотиды РНК включают урацил, цитозин, аденин, гуанин, а ДНК – вместо первого пиримидинового основания имеют его метилированное производное – тимин; 2) как уже описано выше, мононуклеотиды РНК и ДНК содержат разные пентозы; 3) иногда, кроме известных пуринов и пиримидинов, регистрируются их модифицированные аналоги (метилгуанин, метиладенин, дигидроурацил, метилцитозин, и т. д.) – названные минорными основаниями; разные полинуклеотиды отличаются их содержанием; 4) РНК обычно представляют одну цепь, а ДНК эукариот – это двойная нить; 5) молекулярная масса любых РНК меньше, чем ДНК; 6) содержание в клетках ДНК – величина постоянная, а РНК – лабильная; 7) отличаются нуклеиновые кислоты функциями и локализацией.
Виды РНК
Среди РНК различают: информационную, рибосомальную, транспортную, регуляторную, гетерогенно ядерную и др. На долю транспортной РНК (тРНК) приходится 10-15% всех РНК клетки, молекулярная масса не превышает 30 тыс. Д, ее нить состоит из 70-95 мононуклеотидов, причем в ее молекулу включено до 8-19% минорных оснований (дигидрацил, инозин, тиоурацил и т. д.). Необычность их структуры мешает нуклеазам разрушать тРНК, что делает их долгожителями. Оба конца нити тРНК у всех их представителей имеют одинаковое строение: в начале цепи расположен ГМФ, а заканчивается следующим тринуклеотидом: ЦЦА. Вторичная структура представлена в форме клеверного листа, образованной в виде шпилькообразных структур, за счет комплементарности (схема). Сближенные концы цепи тРНК называют акцепторным участком, сюда крепится транспортируемая аминокислота, противоположная петля включает антикодон – триплет, с помощью которого узнается соответствующее (комплементарное) звено на иРНК. Исходя из того, что смысловых кодонов 61, следовательно, и разных тРНК в клетке не менее 60. Еще две петли служат для взаимодействия с рибосомой и аминоацил-тРНК-синтетазой. Цепи различных тРНК укладываются в компактные третичные структуры; за счет стекинг - взаимодействий петли сближаются, принимая форму, напоминающую латинскую букву «L». Митохондриальные тРНК несколько короче (до 65 нуклеотидов), петли также меньше, пространственная структура не установлена.
Схема 3.2.3.1.1. Универсальная укладка полинуклеотидной цепи тРНК.
Функции т-РНК: а) акцепторная – способность ковалентно связываться с аминокислотой, превращаясь в аминоацил
Большая часть цитоплазматических РНК (80-85%) составляют рибосомальные (рРНК), составляющие основу соответсвующих органелл, локализирующихся и в цитозоле, и в митохондриях. Выделяют несколько рРНК (5S, 16-18S, 23-29S) с различными ММ (от 40.000 до 700.000 Д), с неодинаковым числом мононуклеотидов (120-3000). Первичная структура включает минорные основания, хотя и в меньшем количестве, чем у тРНК.
Дальнейшее усложнение организации приводит к спирализации (с помощью водородных, координационных связей), возникновению шпилек с дефектами комплементарности (пиримидин - пиримидин). С помощью шпилек рРНК связываются со своими белками, обычно гистонами, образуя в целом рибосомы, предварительно плотно упаковавшись в пространстве (третичная структура), принимая разные формы (У или V). Функция рибосом – создать каркас, где бы происходило взаимодействие иРНК и тРНК и рост полипептидной цепи. Матричная (информационная) РНК (иРНК) резко отличается от других вариантов полирибонуклеотидов по структуре. На ее долю приходится от 2 до 6% от числа подобных соединений, молекулярная масса колеблется от 25000 до миллиона Д; может включать от сотни до 3000 мононуклеидов. В первичной структуре отсутствуют минорные основания, с одной стороны цепь заканчивается
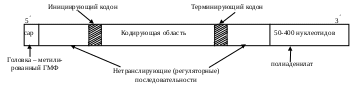
Схема первичной структуры и-РНК.
одинаковыми кирпичиками (50-400 моноаденилатов), подобный неспецифический хвост мешает разрушению иРНК нуклеазами. С другого конца имеет иРНК модифицированный ГМФ (сар-участок), с помощью которого крепится к рибосоме (схема). В середине молекулы между инициирующим и терминирующим кодонами располагается собственно кодирующая область – представляющая чередование информативных и неинформативных участков. Вторичная и третичная структуры индивидуальны, но в итоге имеется несколько шпилек, куда крепятся факторы инициации и терминации, а молекула обязательно связывается со специфическим белком, образуя информосому, сближается с рибосомой, где служит матрицей в синтезе соответствующей полипептидной цепи.
Варианты ДНК
Дезоксирибонуклеиновые кислоты (ДНК) обычно локализуются в ядре и митохондриях, что и лежит в основе их деления.
Принципы строения их молекул однотипны. Как любой биополимер имеют несколько уровней организации: первичную – двойную полинуклеотидную нить, состоящую из дезоксирибонуклеозидфосфатов, связанных фосфодиэфирными связями; вторичную – полученную в результате спирализации; третичную – суперспиралированную структуру. Но так как они сильно отличаются по массе, то и в деталях их строения имеется много отличий.
М
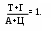
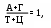
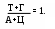 итохондриальную ДНК иногда называют 25–ой хромосомой, ее значения не превышают 5% от общего количества ДНК; в каждой органелле содержится 5-10 ее копий. Это довольно маленький биополимер: его первичную структуру составляют 16569 пар мононуклеотидов. Облигатной чертой ДНК эукарит является спаренность нитей за счет комплементарности. Первым эту особенность заметил Chargaff в 1949 г. Он обратил внимание, что часть и пиримидиновых, и пуриновых оснований включает или кето- или аминогруппы, между которыми за счет наличия водорода в последней могут образовываться водородные связи. Его первый постулат: молярная доля пуринов равна молярной доле пиримидинов и это логично – две нити будут параллельны только тогда, если против крупной структуры (пурина) встанет маленькая (пиримидин). Если подобная ситуация создастся между двумя пуринами – им будет тесно, а пиримидинам – просторно, что не будет способствовать возникновению водородных связей. Во-вторых, для этого необходимо, чтобы против кетогруппы стояла аминогруппа, то есть их число должно быть равным
итохондриальную ДНК иногда называют 25–ой хромосомой, ее значения не превышают 5% от общего количества ДНК; в каждой органелле содержится 5-10 ее копий. Это довольно маленький биополимер: его первичную структуру составляют 16569 пар мононуклеотидов. Облигатной чертой ДНК эукарит является спаренность нитей за счет комплементарности. Первым эту особенность заметил Chargaff в 1949 г. Он обратил внимание, что часть и пиримидиновых, и пуриновых оснований включает или кето- или аминогруппы, между которыми за счет наличия водорода в последней могут образовываться водородные связи. Его первый постулат: молярная доля пуринов равна молярной доле пиримидинов и это логично – две нити будут параллельны только тогда, если против крупной структуры (пурина) встанет маленькая (пиримидин). Если подобная ситуация создастся между двумя пуринами – им будет тесно, а пиримидинам – просторно, что не будет способствовать возникновению водородных связей. Во-вторых, для этого необходимо, чтобы против кетогруппы стояла аминогруппа, то есть их число должно быть равнымИсходя из выше сказанного, третьим положением и будет: А = Т, а Г = Ц (схема).
В целом, первичная структура ДНК представляет нить, образованную фосфодиэфирными связями между мононуклеотидами одной цепи и водородными между противоположными. (см. Приложение, схема 2). Кроме того, рядом лежащие гидрофобные ароматические основания за счет различных по плотности электронных облаков способны взаимодействовать друг с другом, создавая так называемые стекинг-взаимодействия, а фосфаты, являясь заряженными группами, вступают в ионные связи с различными катионами (металлов, основными белками, полиаминами).
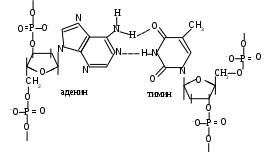
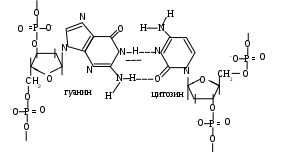
Схема. Водородные связи между основаниями в ДНК.
Характеризуя митохондриальную ДНК следует заметить, что в первичной структуре отсутствуют интроны, в ее
кодогенах (триплетах - комплементарных смысловым кодонам иРНК) нередки отклонения от генетического кода. Вторичная структура - хорошо спирализованное образование. Третичная - представляет кольцевой дуплекс, то есть концы сдвоенной нити сближены и соединены между собой. ДНК митохондрий содержит информацию не обо всех белках этих органоидов (см. «Патохимия наследственных болезней» С. 18).
Самая большая молекула, содержащая в клетках, это ядерная ДНК. Если ее вытянуть до первичной структуры, длина составит около 8 см (!). Этот уровень организации включает различные гены (транскриптоны, регуляционные и т.д.), палиндромы. Последние представляют фрагменты нитей, одна половина которой комплементарна второй, при их сближении друг с другом за счет возникновения водородных связей образуются шпильки, к которым крепятся различные белки (регуляторные, каталитические).
Первичная структура включает и минорные основания, о вероятной роли которых было написано выше.
Как замечено раньше, нить с одного конца фосфорилирована (5’ –конец), на другом расположена свободная 3’–ОН группа дезоксирибозы (3’ – конец). Когда две нити связаны одной ДНК, то они антипараллельны - каждый конец цепи имеет 3’ и 5’ концы.
Ядерная ДНК хорошо спирализована - это правозакрученная спираль, шаг которой равен 3,4 нм, диаметр составляет 2 нм, 1 виток включает 10 мононуклеотидов и полюс шпильки. Подобная конформация известна как В-форма и обычна для клетки. Но молекула ДНК может менять свою упаковку в зависимости от условий: быть более сплющенный (А-форма), зигзагообразный (Z-форма).
Важным свойством двойной спирали ДНК является ее способность изгибаться. Молекула ее в миллион раз длиннее, чем ядро или клетка и чтобы поместиться в них, она должна быть гибкой, что приводит к суперспирализации , т.е. третичная структура представляет дальнейшее закручивание двойной нити вокруг положительно заряженных белков - гистонов. Последние образуют октамерный (восемь, т.е 4 пары Н2а, Н2в, Н3, Н4 гистонов) протеиновый комплекс, называемый нуклеосомой, вокруг которого молекула ДНК совершает два оборота (схема).
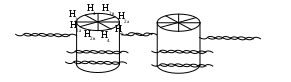
Схема. Модель упаковки ДНК
Участок ДНК, расположенный между рядом лежащими нуклеосомами и включающий 30-40 пар мононуклеотидов (в отличие от 200 пар, окружающих катушку), - называют линкерным. Считают, что он связан с Н1-гистонами. Подобная упаковка доводит до стократного уменьшения молекулы. В дальнейшем подобные нити образуют петли, что увеличивает компактность; за счет дополнительной спирализации и обеспечивается 10000-кратная конденсация ДНК.
Физико-химические и биологические свойства нуклеиновых кислот
Значительное содержание фосфора (имеющий значительный атомный вес), плотная, особенно у ДНК упаковка обуславливают высокую плотность, достигающую 1,7 у ядерной ДНК. За счет спирализации, а у ДНК - удвоения нитей поверхности молекул полинуклеотидов образованы полярными участками пентоз и фосфатов, что, во-первых, обеспечивает им хорошую растворимость, а во-вторых - создает кислую среду. Комплексирование же нуклеиновых кислот (полианионов) с белками (поликатионами) снижает величину заряда нуклеопротеида, тем самым уменьшает его способность растворяться.
Чем больше их молекулярная масса, тем выше их плотность, вязкость растворов, измеряются осмотическое и онкотическое давление. Как молекулы белков, которые под действием факторов, разрывающих добавочные связи (Приложение, схема 1), денатурируют, так и в нуклеиновых кислотах под влиянием повышенной температуры (85 – 1000), лучевой энергии, ультразвука, химических реагентов, изменяющих рН, происходит подобное явление (денатурация). Однако, если действие фактора кратковременное и неглубокое, то вполне вероятно восстановление поврежденных связей (гибридизация).
Самая крупная молекула ядерной ДНК - хрупкая, довольно легко ломается, например, под действием ультразвука.
Среди биологических свойств ДНК следует выделить:
-
Репликацию - её способность работать в качестве матрицы, т.е. на материнской нити синтезируется новая ядерная ДНК. -
Транскрипцию - способность участков ДНК служить матрицей в синтезе РНК. -
Обратную транскрипцию - способность фрагментов ДНК образовываться на базе РНК. -
Модификацию - способность ДНК преобразовывать включенные в ее состав ароматические основания путем метилирования, гидрирования. -
Рестрикцию - возможность с помощью различных нуклеаз вычленять отдельные фрагменты из состава ДНК. -
Репарацию - способность восстанавливать поврежденные участки ДНК. -
Интеркаляцию - возможность ароматического основания какого-либо участка ДНК или другого вещества (антибиотика и т.д.) внедриться между двумя соединениями, вначале разорвав соединение, а позднее образовав новые стекинг-взаимодействия.
Катаболическая фаза обмена нуклеотидов