ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 09.12.2023
Просмотров: 308
Скачиваний: 4
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
Таким образом, структурная организация восприятия первичной локализованной боли (эпикритической боли) представляется следующим образом.
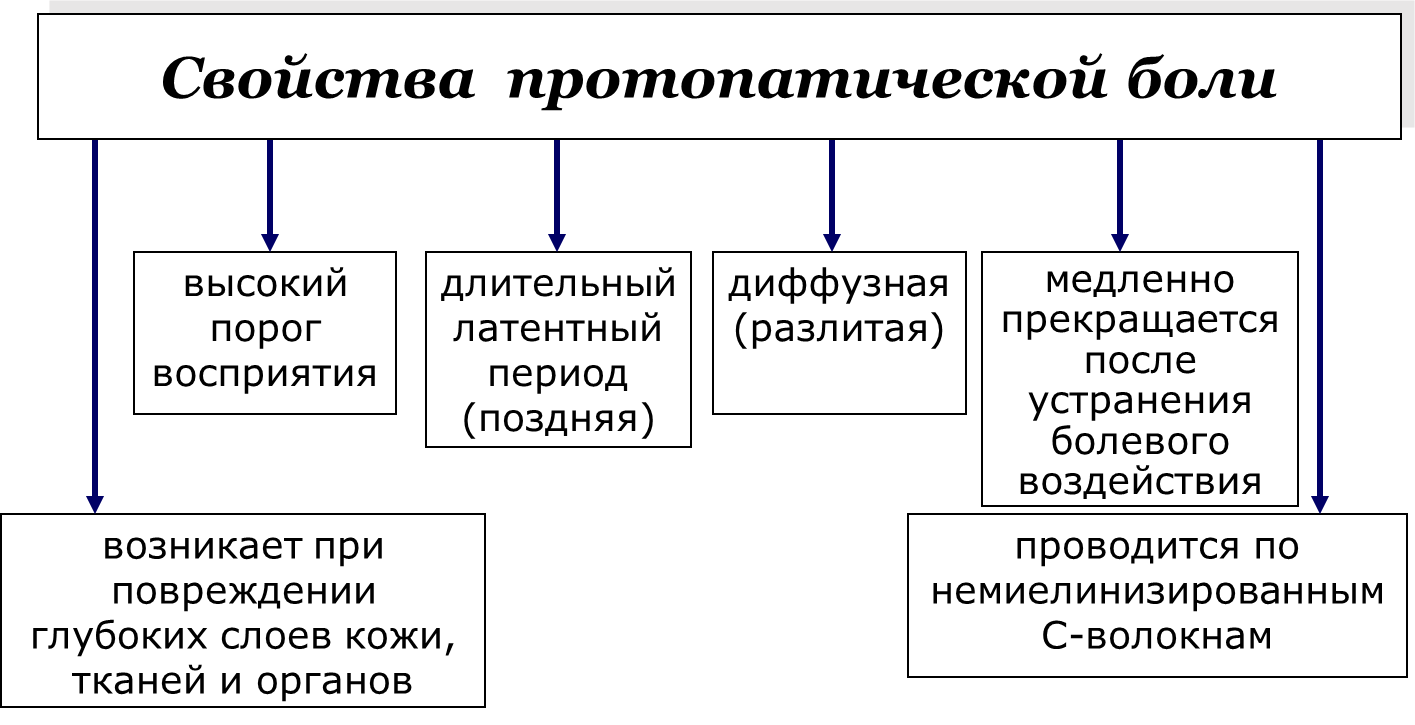
Рис. 9. Характеристики поверхностной протопатической (поздней) боли.
Нейрон 1-го уровня – спинномозговые ганглии (узлы);
нейрон 2-го уровня – задние рога спинного мозга;
нейрон 3-го уровня – таламус (вентробазальные ядра);
нейрон 4-го уровня – соматосенсорные зоны коры.
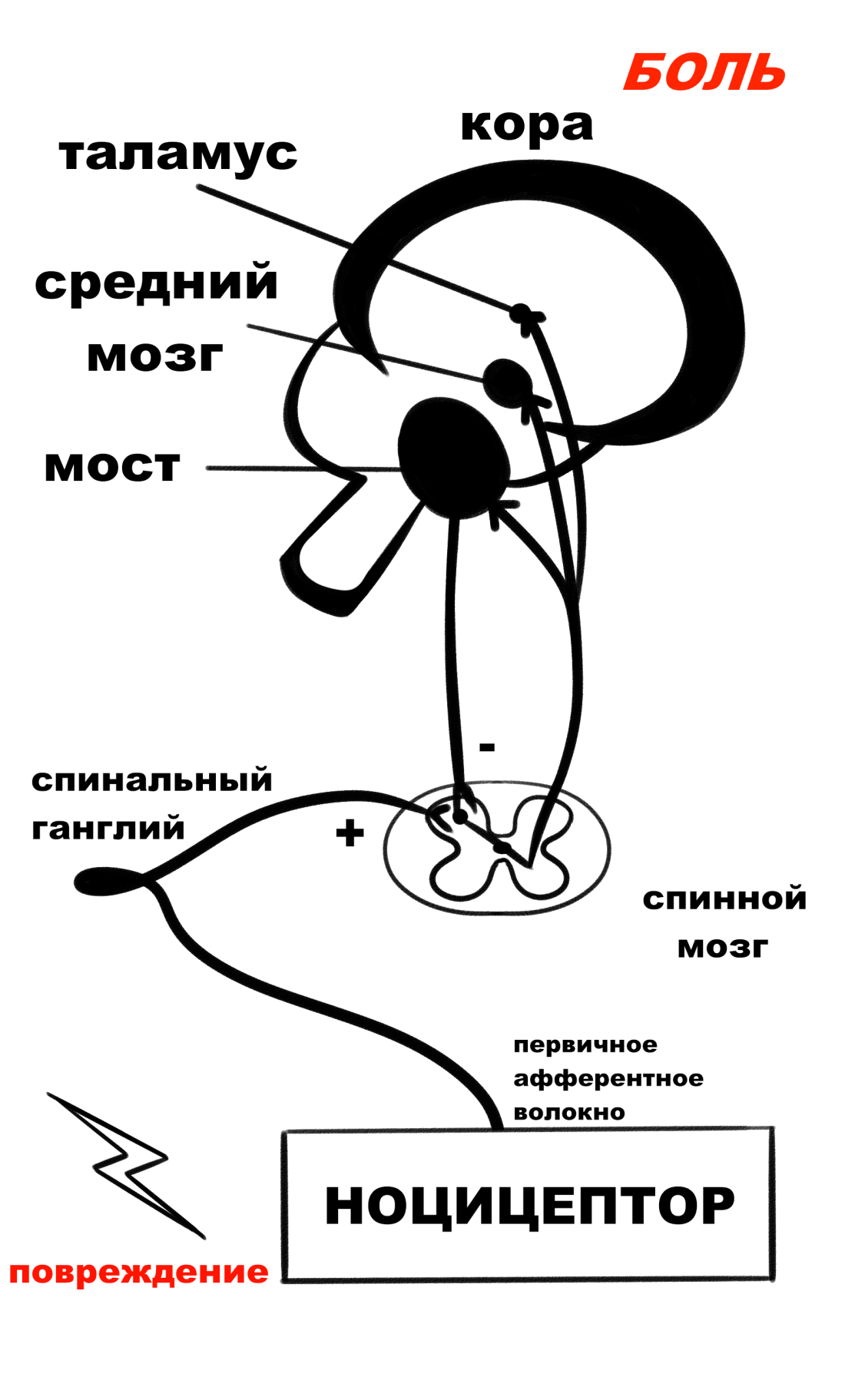
Рис. 10. Схема взаимодействия структур восприятия первичной локализованной боли.
Медиаторы боли
На сегодняшний день известно несколько десятков нейрохимических соединений, которые вовлечены в процессы возбуждения и торможения болевых нейронов в ЦНС. Нейрогормоны, нейромедиаторы и нейромодуляторы могут представлять собой низкомолекулярные (например, аминокислоты) и более сложные высокомолекулярные соединения (субстанция Р, нейрокинин А, кальцитонин ген-связанный пептид и др.). Так, возбуждающие аминокислоты – ВАК (глутамат, аспартат) способны опосредовать и играть значимую роль в проведении ноцицепции. Глутамат содержится более чем в половине нейронов спинальных ганглиев и высвобождается под действием ноцицептивных импульсов. ВАК взаимодействуют с несколькими подтипами глутаматных рецепторов. Это, прежде всего, ионотропные рецепторы: NMDA–рецепторы(N–метил–D–аспартат) и АМРА–рецепторы(α–амино–3–гидрокси–5–метил–4–изоксазол–пропионовой кислоты), а также метаболотропные глутаматные рецепторы(mGluR). Активация этих рецепторов приводит к значительному увеличению поступления ионов Са2+ внутрь клетки, что способствует её дальнейшей физиологической модуляции. Гипералгезия в этом случае будет возникать в результате формирования очагов стойкой гипервозбудимости нейронов. Сенситизация этих болевых нейронов может сохраняться несколько часов или дней после прекращения поступления ноцицептивных импульсов из повреждённого участка ткани на периферии. Таким образом, если гиперактивация ноцицептивных нейронов сформировалась, то постоянного дополнительного поступления сигналов из места повреждения для поддержания этого очага не требуется.
Экспрессия ранних, немедленно реагирующих генов, таких как
c–fos, c–jun, junB и некоторых других, способствует длительному и существенному повышению возбудимости болевых нейронов. Так, например, имеются данные о положительной корреляции между количеством fos-позитивных нейронов и степенью боли. Также, важное значение в механизмах активации протоонкогенов отводят ионам Са2+. При повышении концентрации ионов Са2+ в цитозоле, вследствие усиленного их входа через регулируемые NMDA-рецепторами Са–каналы, происходит экспрессия с–fos, с–jun, белковые продукты которых участвуют в регуляции долговременной возбудимости мембраны клетки.
В последнее время всё больше данных свидетельствует об участии оксида азота(NO) в механизмах сенситизации ноцицептивных нейронов. Эта молекула выполняет функцию нетипичного внесинаптического медиатора в мозге. Благодаря своим малым размерам и отсутствию заряда NO может проникать через плазмалемму и участвовать в межклеточной передаче сигнала, функционально соединяя пре- и постсинаптические нейроны. NO образуется из L-аргинина в нейронах, содержащих фермент NO-синтетазу. NMDA-индуцируемое возбуждение способствует выделению NO из клеток и взаимодействию с пресинаптическими терминалями С-афферентных волокон. В терминалях этих афферентов в этом случае повышается выброс возбуждающей аминокислоты глутамата и нейрокининов. Доказано участие оксида азота в развитии воспалительных процессов. Так, например, местное введение ингибиторов NО-синтазы в сустав эффективно блокирует ноцицептивную передачу и воспалительную реакцию. Эти данные подтверждают тот факт, что оксид азота в избытке образуется при артритах.
Одними из наиболее сильных болевых модуляторовявляются кинины. Они быстро образуются при повреждении ткани и вызывают большинство эффектов, наблюдаемых при воспалении: вазодилатацию, увеличение сосудистой проницаемости, экстравазацию плазмы, миграцию клеток, боль и гипералгезию. Кинины активируют С-афференты, что приводит к нейрогенному воспалению за счет выброса из нервных терминалей субстанции Р, кальцитонин ген-связаннного пептида и других нейромедиаторов. Одним из наиболее активных кининов-медиаторов боли является брадикинин, который представляет собой пептид, состоящий из 9 аминокислот. Он образуется на месте повреждения благодаря действию ферментов. Фактор XII (Хагемана) проникает в поврежденные ткани и превращается в них в активную форму. Затем, воздействуя на прекалликреин, переводит его в калликреин, а последний конвертирует белок-предшественник кининоген в брадикинин. Брадикинин способен вызывать у человека болевые ощущения в концентрациях, эквивалентных обнаруживаемым в поврежденных тканях. Рецепция брадикинина происходит на чувствительных волокнах и в задних рогах спинного мозга. Наряду с этим брадикинин может повышать сосудистую проницаемость, стимулировать хемотаксис лейкоцитов и сенсибилизировать ноцицепторы.
Прямой возбуждающий эффект брадикинина на сенсорные нервные терминали связан с активацией мембранной фосфолипазы С и может быть опосредован B2-рецепторами. Непрямое возбуждающее действие брадикинина на окончания нервных волокон обусловлено его воздействием на различные тканевые элементы (эндотелиальные клетки, фибробласты, тучные клетки, макрофаги и нейтрофилы) и стимулированием образования в них медиаторов воспаления, которые, взаимодействуя с соответствующими рецепторами на нервных окончаниях, активируют мембранную аденилатциклазу. В свою очередь, аденилатциклаза и фосфолипаза С стимулируют образование ферментов, фосфорилирующих белки ионных каналов. Результатом фосфорилирования белков ионных каналов является изменение проницаемости мембраны для ионов, что отражается на возбудимости нервных окончаний и способности генерировать нервные импульсы. Брадикинин, действуя через В2-рецепторы, стимулирует образование арахидоновой кислоты с последующим образованием простагландинов, простациклинов, тромбоксанов и лейкотриенов. Эти вещества, обладая выраженным самостоятельным альгогенным действием, в свою очередь, потенциируют способность гистамина, серотонина и брадикинина сенситизировать нервные окончания. В результате этого из немиелинизированных С-волокон усиливается выброс тахикининов (субстанции Р и нейрокинина А), которые, увеличивая сосудистую проницаемость, еще больше повышают локальную концентрацию медиаторов воспаления.
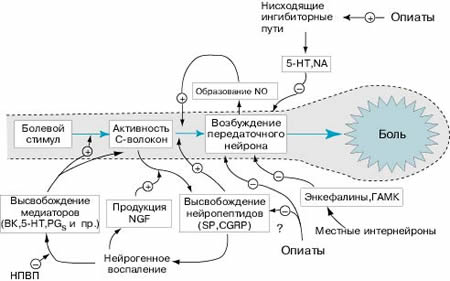
Рис. 11. Механизмы регуляции ноцицептивного пути (Ивашкин В.Т., Шульпекова Ю.О., 2002).
Примечание: NGF – фактор роста нервов, BK – брадикинин, 5-НТ – 5-гидрокситриптамин (серотонин), PGs – простагландины, NA – норадреналин, SP – субстанция Р, CGRP – кальцитонин ген-связанный пептид.
Нейрогуморальная регуляция болевой чувствительности осуществляется на разных уровнях и представлена большим многообразием факторов, которые представлены на рис. 11. Ноцицептивный стимул воспринимается болевыми чувствительными волокнами, передающими сигналы передаточным нейронам спиноталамического тракта. Далее по таламокортикальным волокнам импульс достигает коры головного мозга, где формируется восприятие боли. Передача болевого возбуждения с периферии на передаточные нейроны спиноталамического тракта облегчается посредством NO, вещества P (SP) и кальцитонин ген-связанного пептида (CGRP). Медиаторами нисходящих цереброспинальных антиноцицептивных импульсов могут выступать серотонин (5-HT) и норадреналин (NA). Медиаторами антиноцицептивных импульсов от нейронов желатинозной субстанции (SG-нейронов) являются энкефалины, ГАМК. При нейрогенном воспалении наблюдается избыточное и длительное высвобождение нейропептидов SP, CGRP из С-волокон, поддерживаемое такими воспалительными веществами, как брадикинин (BK), 5-HT, простагландины (PGs) и фактор роста нервов (NGF). Применение нестероидных противовоспалительных препаратов позволяет уменьшить продукцию воспалительных медиаторов. Опиаты снижают болевую чувствительность посредством активации нисходящих антиноцицептивных сигналов и угнетения передаточных нейронов спиноталамического тракта.
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ АНТИНОЦИЦЕПТИВНОЙ СИСТЕМЫ
Формирование боли как психофизиологического состояния имеет в основе интегративную деятельность мозга. Характер и интенсивность специфического болевого ощущения обеспечиваются динамическим взаимодействием эволюционно сформированных ноцицептивной и антиноцицептивной систем. В физиологических условиях эти 2 системы согласованно функционируют. Воздействие альгогенов стимулирует ноцицептивную передачу, что в свою очередь способствует активации антиноцицептивной системы. Это приводит к лимитированию болевой афферентации, восприятия боли, метаболических, вегетативных, поведенческих реакций и, в целом, способствует более быстрому и эффективному восстановлению нарушенных структур и функций.
Антиноцицептивная система (АНЦС) имеет свои центры, пути, нейротрансмиттеры и нейромодуляторы. В ней можно выделить собственно нервный аппарат и нейрохимические механизмы.
Нервный аппарат антиноцицептивной системы представлен следующими образованиями.
-
Центральное серое околоводопроводное вещество (ЦСОВ). -
Ядра шва (особенно большое – БЯШ). -
Ядра покрышки среднего мозга. -
Сегментарный аппарат спинного мозга (интернейроны желатинозной субстанции – SG).
Главенствующее положение в иерархии антиноцицептивной системы в головном мозге принадлежит центральному серому околоводопроводному веществу (ЦСОВ) среднего мозга. Эта его роль обусловлена анатомической близостью к структурам, в которые направляются чувствительные волокна восходящих ноцицептивных проводников. Описано также наличие проекций спинальных афферентов и восходящих проекций ретикулярной формации (РФ) в ЦСОВ. В эксперименте впервые был выявлен феномен селективной анестезии при электрораздражении, преимущественно вентральных отделов околоводопроводного вещества. ЦСОВ не имеет прямых связей со спинным мозгом, все его двигательные терминали оканчиваются на нейронах группы ядер РФ и БЯШ.
Другой областью эндогенной болеутоляющей системы являются ядра шва ствола и среднего мозга. Они тесно связаны с ЦСОВ, а аксоны их клеток проецируются как в восходящем, так и в нисходящем направлениях. Нисходящие волокна из этих ядер, особенно из большого ядра шва (БЯШ), проходят в дорсолатеральном канатике и прослеживаются в заднем роге спинного мозга.
Стимуляция ЦСОВ и ядер шва тормозит активность нейронов заднего рога спинного мозга, связанных с ноцицептивной афферентацией. Особенно отчетливо изменяется активность нейронов спиноталамического тракта. Раздражение ЦСОВ, угнетающее активность спиноталамических нейронов спинного мозга, одновременно активирует клетки БЯШ. Кроме того, стимуляция ЦСОВ вызывает активацию нейронов желатинозной субстанции, которая участвует в пресинаптическом торможении ноцицептивной информации.
Наряду с нисходящей системой ядер шва существует система нисходящих связей ядер ретикулярной формации ствола головного мозга.
Раздражение этой области вызывает отчётливое обезболивание и угнетает ответы нейронов заднего рога спинного мозга на ноцицептивные стимулы. По некоторым данным, стимуляция ретикулярных ядер ствола в значительно большей степени, чем раздражение ядер шва, угнетает передачу ноцицептивной информации в спинном мозге. В отличие от волокон, нисходящих из ядер шва, ретикулоспинальные пути оканчиваются не только в I—V слоях заднего рога, но и в боковом и передних рогах, что имеет существенное значение для сопряженной регуляции вегетативной и моторной деятельности при стимуляционной анальгезии. В настоящее время значительное внимание уделяется изучению роли в регуляции болевой чувствительности латерального ретикулярного ядра, раздражение которого сопровождается отчетливой анальгезией, что реализуется через дорсолатеральные и вентролатеральные канатики. Некоторые нейроны латерального ядра выполняют функцию релейного звена в системе ЦСОВ — ядра шва — спинной мозг и имеют морфологические прямые проекции из ЦСОВ и ядер шва.
Исследователями недавно получены данные о том, что стимуляция ЦСОВ тормозит активность ноцицептивных клеток в медиальном таламусе. Установлено также, что некоторые клетки ЦСОВ увеличивают свою активность при периферическом ноцицептивном раздражении, что совпадает с моментом активации на ЭЭГ в коре больших полушарий.
ЦСОВ имеет тесные связи с лимбической системой и интраламинарными ядрами таламуса. ЦСОВ получает афференты из гипоталамуса и миндалины и, в свою очередь, посылает эфферентные волокна в парасимпатические центры продолговатого мозга.
Помимо восходящих проекций из ЦСОВ существуют такие восходящие проекции из ядер шва, которые могут оказывать модулирующее влияние на ноцицептивные реакции не только на уровне спинного мозга, но и на выше лежащих уровнях, в частности в таламусе. Наличие прямых проекций от ядер шва обнаружено также к парафасцикулярному и вентробазальному комплексам таламуса, хвостатому ядру, латеральной перегородке, гиппокампу и мозжечку. Установлены двусторонние моносинаптические связи БЯШ с гипоталамусом и гиппокампом.