ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 27.11.2019
Просмотров: 4034
Скачиваний: 98
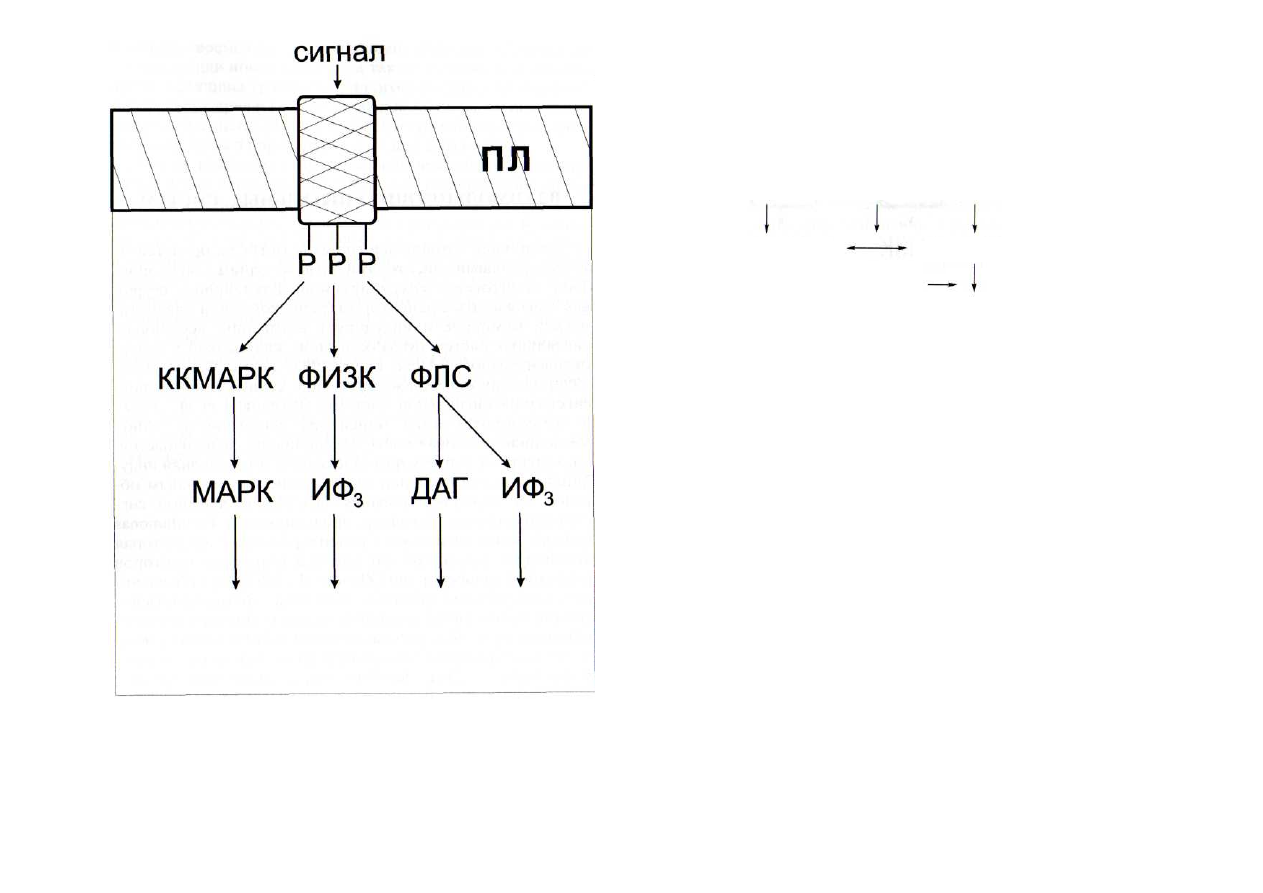
Рис. 40. Дивергенция сигнальных путей, обусловленная несколь-
кими сайтами фосфорилирования у одной молекулы рецептор-
ной протеинкиназы [Cohen, 2000]
ДАГ - диацилглицерол; ИФ
3
-
инозитолтрисфосфат; ККМАРК -
киназа киназы МАР-киназы; МАРК - митогенактивируемая протеин-
киназа; ПЛ - плазмалемма; Р - остатки фосфорной кислоты; ФИЗК -
фосфатидилинозитол-3-киназа; ФЛС - фосфолипаза С
[Delledone et al.,
1998], так и интеграции некоторых из них
[Cheng et al.,
1998; и др.].
Одним из наиболее простых случаев взаимодействия
сигнальных систем может считаться взаимопревращение
"стартовых" фосфолипидов, принадлежащих двум различ-
ным сигнальным системам - кальциевой и фосфатидатной
[Munnik et al., 1998; Munnik, 2001]:
ФЛС
ДАГ-киназа
Диацилглицерол-
пирофосфат
Найдено образование из фосфатидной кислоты диацил-
глицеролпирофосфата [Van der Luit et al., 2000], роль кото-
рого в сигнальных процессах может быть двоякой: снижение
содержания основного сигнального соединения фосфатида-
та и активация специфической протеинкиназы. Возмож-
ность активации протеинкиназ диацил-глицеролом, лизо-
фосфатидами и фосфатидной кислотой уже отмечалась в
разделах, посвященных кальциевой, липоксигеназной и
фосфатидатной сигнальным системам.
Отмечено взаимодействие сигнальных систем на уровне
факторов регуляции транскрипции, имеющих много мест
фосфорилирования, которые могут обслуживаться проте-
инкиназами, активируемыми разными сигнальными систе-
мами [Hill, Treisman, 1995], что еще более выражено у оли-
гомерных активных форм факторов регуляции транскрип-
ции, по сравнению с неактивными мономерными [Cohen,
2000] (рис. 41).
К настоящему времени накопилось немало фактов, сви-
детельствующих о возможности модулирования (активации
или ингибировании) одних сигнальных систем с помощью
промежуточных продуктов (вторичных посредников) дру-
гих. В циклоаденилатной системе таким сигнальным интер-
медиатом является цАМФ, в фосфатидатной - фосфатид-
ная кислота, в МАР-киназной - различные протеинкиназы,
в кальциевой - инозиттрисфосфат и инозиттетракисфос-
фат, диацилглицерин и ионы кальция, в липоксигеназной -
Диацилглицерол
Фосфатидат
Фосфатидат-киназа
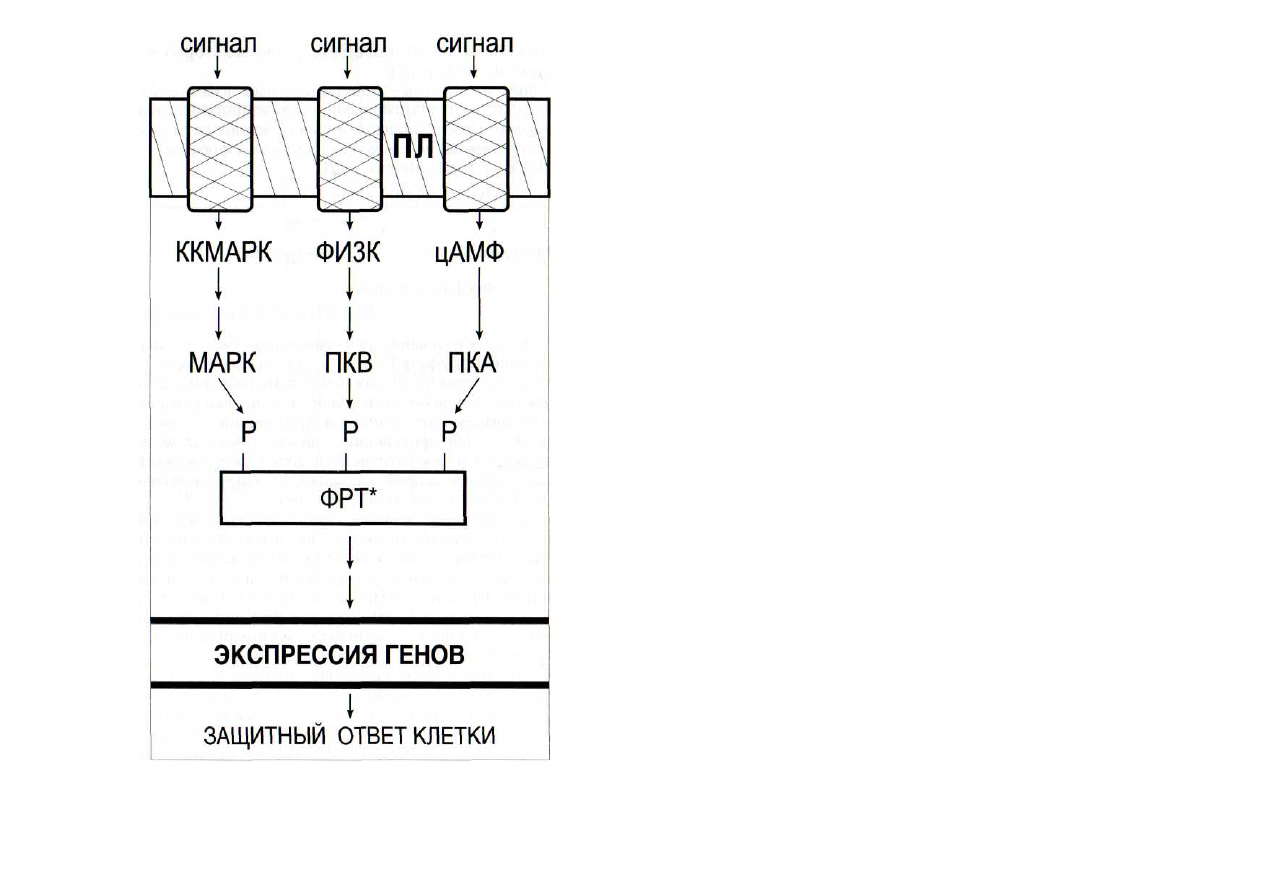
Рис. 41. Конвергенция сигнальных систем [Cohen, 2000]
ПКА и ПКВ - протеинкиназы А и В; ФРТ - фактор регуляции
транскрипции. Остальные обозначения - см. рис. 40
полиеновые жирные кислоты, их гидроперокси-, гидрокси-,
эпокси-, кето-, циклические и другие производные, в
НАДФН-оксидазной - активные формы кислорода (напри-
мер, супероксидный анионрадикал и перекись водорода) и
салицилат, в NO-синтазной - окись азота, цГМФ, цАДФ-ри-
боза и салицилат. Если активация (или инактивация) фер-
ментов сигнальных систем или белков ионных каналов осу-
ществляется с помощью фосфорилирования или дефосфо-
рилирования, то в качестве сигнальных интермедиатов мо-
гут выступать, соответственно, протеинкиназы или проте-
инфосфатазы.
К числу сигнальных интермедиатов кальциевой сиг-
нальной системы растений относятся различные изофор-
мы кальмодулина, причем одни из них активируют, а дру-
гие при той же концентрации инактивируют определен-
ную Са
2+
-
кальмодулинзависимую (например, NO-синтаз-
ную) реакцию. В зависимости от концентрации той или
иной изоформы могут активироваться или ингибировать-
ся ферменты различных сигнальных систем. Неодинако-
вая интенсивность экспрессии изоформ кальмодулина в
зависимости от условий (при действии различных типов
элиситоров) может определять преимущественное вклю-
чение той или иной сигнальной системы [Cho et al., 1998].
Кальмодулин может оказывать неодинаковое влияние не
только на различные сигнальные системы, но и на звенья
одной и той же системы. Например, Са
2+
-
кальмодулин мо-
жет стимулировать не только приходную часть баланса
цАМФ, но и расходную, активируя фосфодиэстеразу
цАМФ [Brown, Newton, 1981]. Разница в степени актива-
ции аденилатциклазы и фосфодиэстеразы (а значит, и со-
держание цАМФ) зависит от концентрации комплекса
Са
2+
—
кальмодулин. Та же закономерность прослеживается
и при анализе влияния ионов кальция и кальмодулина на
активность протеинфосфатаз и в связи с этим на степень
фосфорилированности различных белков, в том числе
участников сигнальных систем.
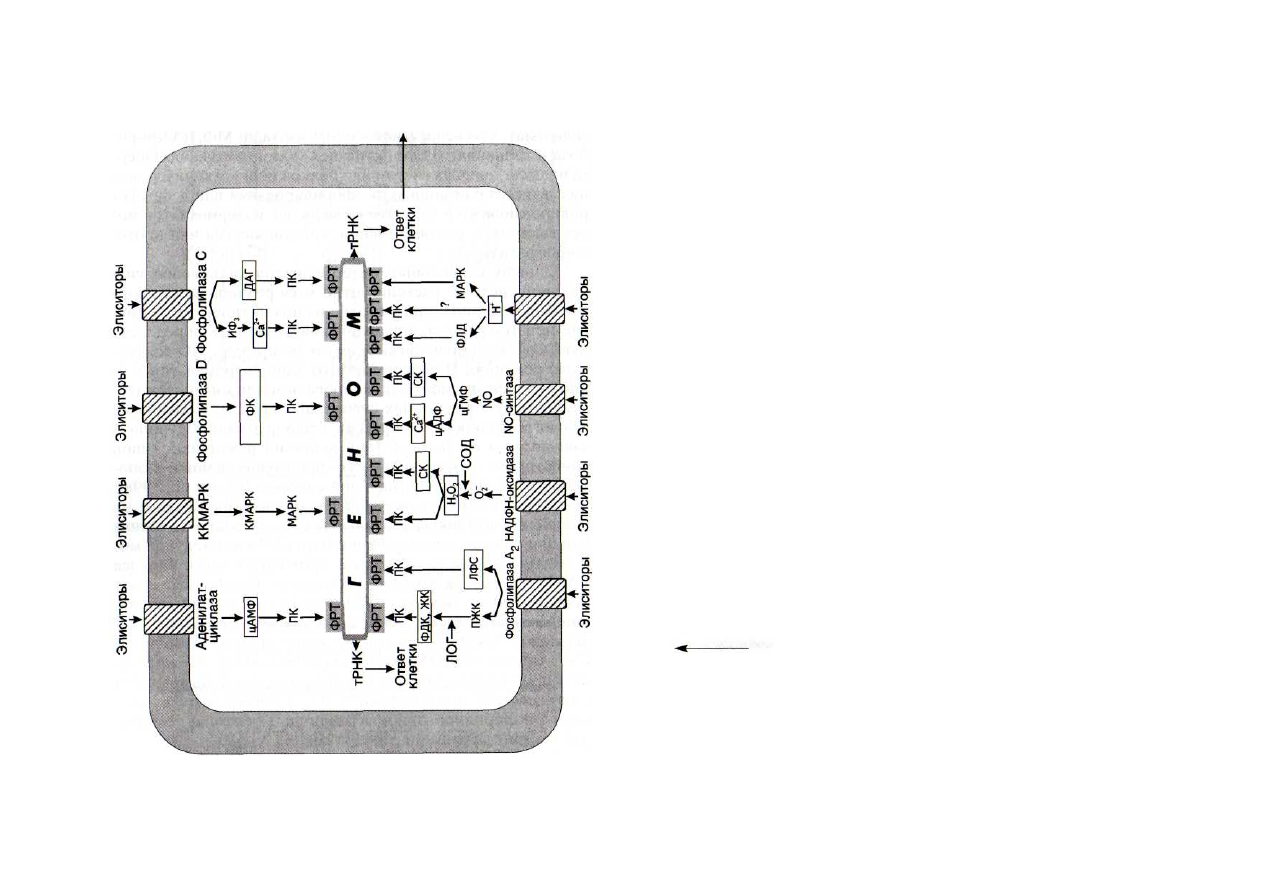
На рис. 42 показаны не только основные участники сиг-
нальных цепей, но и места активации или ингибирования их
основных ферментов. Как и в случае обычных метаболиче-
ских цепей, главным объектом регуляции в сигнальных си-
стемах является фермент первой реакции.
В аденилатциклазной сигнальной системе таким фер-
ментом является аденилатциклаза (рис. 43). Она активиру-
ется относительно низкими концентрациями ионов каль-
ция [Brown, Newton, 1981] и Са
2+
-
кальмодулином, оксиге-
нированными производными полиеновых жирных кислот
[Маеда, Акаике, 1988], но ингибируется относительно вы-
сокими концентрациями Са
2+
[De Bernardi, Brooker, 1996].
Комплекс Са
2+
-
кальмодулин может стимулировать не
только приходную часть баланса цАМФ, но и расходную,
активируя фосфодиэстеразу цАМФ [Brown, Newton, 1981].
Разница в степени активации аденилатциклазы и фосфо-
диэстеразы (а значит, и содержание цАМФ) зависит от
концентрации комплекса Са
2+
-
кальмодулин. Приводятся
также данные об активации фосфодиэстеразы цАМФ с
помощью фосфатидной кислоты [Munnik et al., 1998;
Munnik, 2001].
Как уже отмечалось, интермедиаты липоксигеназной
сигнальной системы ГДК и МеЖК активировали в присут-
ствии цАМФ протеинкиназную активность на 33-48% [Ка-
римова и др., 19996]. Салициловая кислота индуцировала
повышение уровня цАМФ-зависимой фосфорилирован-
ности полипептидов 74, 61, 22 кДа в листьях гороха [Муха-
метчина, 2000]. цАМФ-стимулируемая протеинкиназная ак-
тивность растворимых белков листьев гороха зависела от
концентрации Са
2+
[Каримова и др., 1989; Каримова, Жуков,
1991]. Есть данные об ингибировании моноокисью азота
аденилатциклазы в клетках животных [Watson et al.,
2001].
Функционирование МАР-киназной системы также мо-
жет регулироваться интермедиатами других сигнальных пу-
Рис. 42. Совокупность сигнальных систем клеток растений
ДАГ - диацилглицерол; ЖК - жасмоновая кислота; ИФ
3
-
инози-
толтрисфосфат; ККМАРК - киназа киназы МАР-киназы; КМАРК —
киназа МАР-киназы; ЛОГ - липоксигеназа; ЛФС — лизофосфатиды;
МАРК — митагенактивируемая протеинкиназа; ПЖК — полиеновые
жирные килоты; ПК - протеинкиназы; СК - салициловая кислота;
СОД - супероксиддисмутаза; ФДК - фитодиеновая кислота; ФК - фос-
фатидная кислота; ФЛД - фосфолипаза Д; ФРТ - факторы регуляции
транскрипции; цАДФР - циклическая АДФ-рибоза; цАМФ - цикличе-
ский аденозинмонофосфат; цГМФ — циклический гуанозинмоно-
фосфат
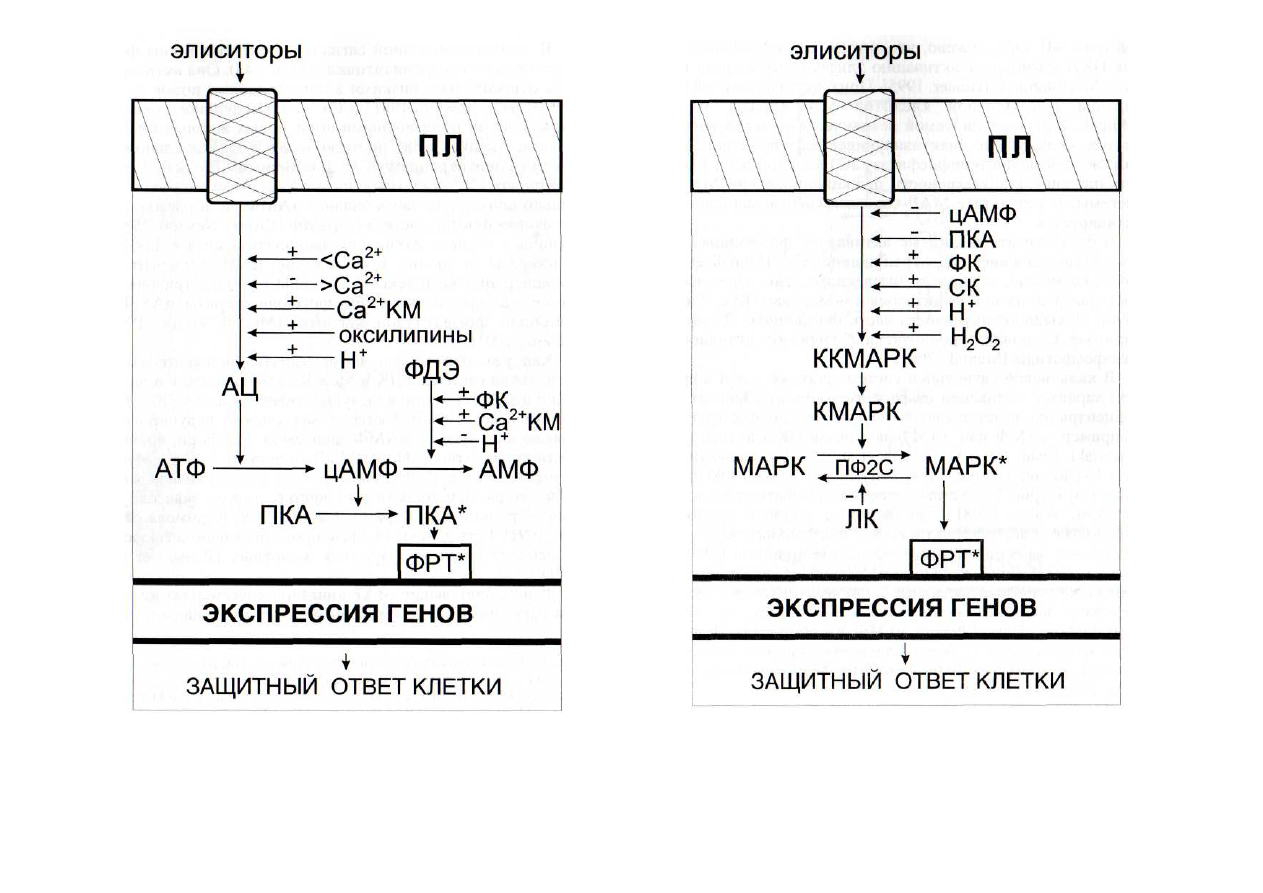
Рис. 43. Регуляция функционирования аденилатциклазной сиг-
нальной системы интермедиатами других сигнальных систем
АЦ - аденилатциклаза; КМ - кальмодулин; ПКА - протеинкиназа
А; ПЛ - плазмалемма; ФДЭ - фосфодиэстераза. > - высокие концен-
трации; < - низкие концентрации. Здесь и на последующих рисунках:
(+) -
активация; (-) - ингибирование. Остальные обозначения - см.
рис. 42
Рис. 44. Регуляция функционирования МАР-киназной сигнальной
системы интермедиатами других сигнальных систем
ЛК - линоленовая кислота; ПКА - протеинкиназа А; ПЛ - плазма-
лемма; ПФ2С - протеинфосфатаза 2С. Остальные обозначения - см.
рис.
42
тей (рис. 44). Обнаружено, что цАМФ или цАМФ-зависи-
мая ПКА ингибирует активацию элиситорами киназы ки-
назы МАР-киназы [Hunter, 1995]. Приводятся данные об ак-
тивации фосфатидной кислотой МАР-киназы WIPK
[Munnik,
2001], индуцируемой механическим повреждением
клеток. Обнаружено инактивирующее действие линолено-
вой кислоты на протеинфосфатазу 2С [Baudouin et al., 1999],
регулирующую интенсивность функционирования МАРК
системы. В результате МАР-киназная сигнальная система
активируется.
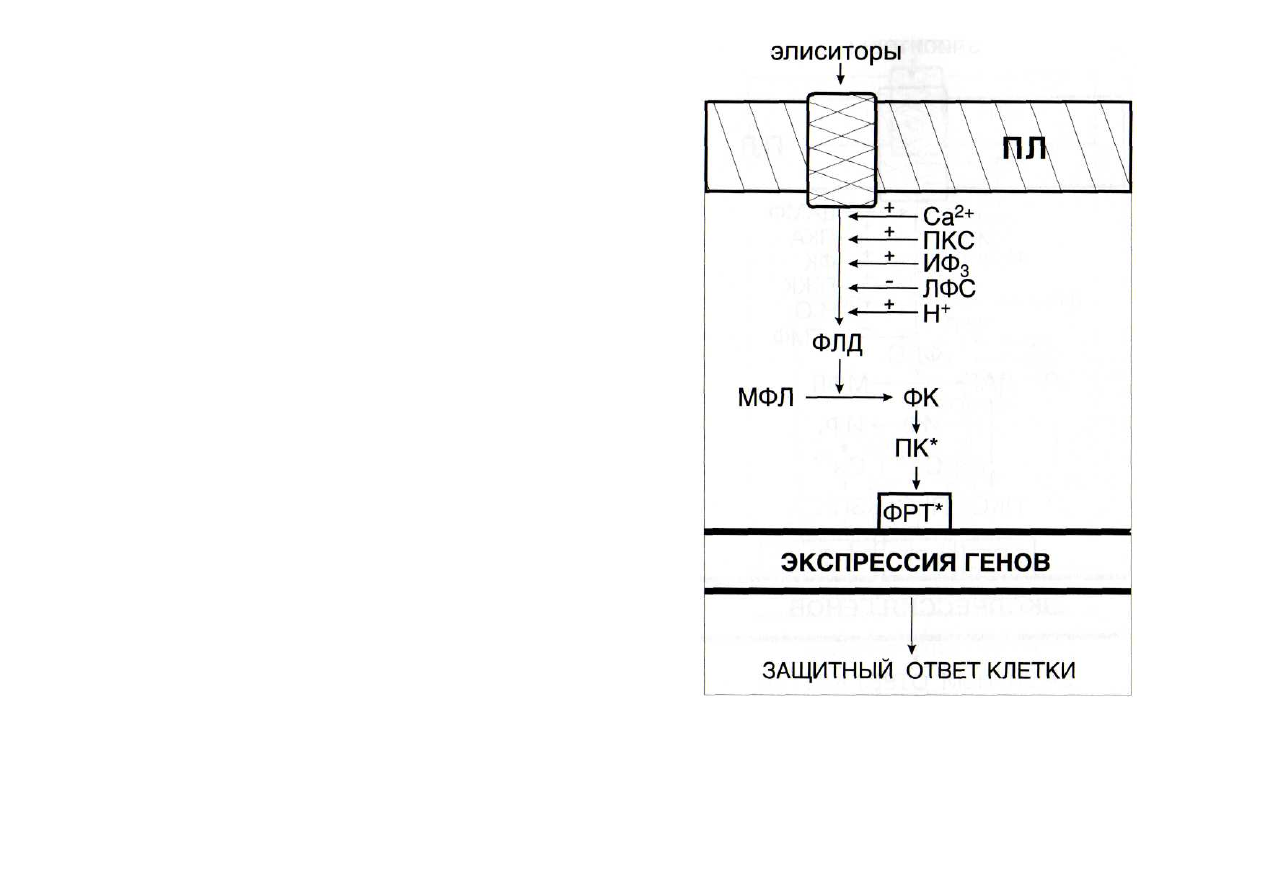
В фосфатидатной системе активирует фосфолипазу Д
(рис. 45) повышение концентрации ионов Са
+
[Munnik et al.,
1995], возможно, вследствие вызванного этим передвиже-
нием фосфолипазы Д из цитозоля к мембранам [Ryu, Wang,
1996].
Повышают активность части фосфолипаз Д проте-
инкиназа С, полифосфоинозитолы, снижают активность
лизофосфатиды [Munnik, 2001].
В кальциевой сигнальной системе (так же как и в дру-
гих) характер активации стартового фермента зависит от
концентраций интермедиатов других сигнальных систем.
Например, цАМФ или цАМФ-зависимая ПКА активирует
[Kurosaki, Nishi, 1993; Volotovsky et al.,
1998] или инактиви-
рует [Авдонин, Ткачук, 1994; Tertyshnikova, Fein,1998] фос-
фолипазу С (рис. 46). Установлено, что фосфатидная кислота
[Ryu, Wang,
1998] и полиеновые жирные кислоты
[Volotovsky et al.,
1998] активируют фосфолипазу С, а пос-
ледние - и кальцийрегулируемые протеинкиназы [Маеда,
Акаике, 1988]. Более сложная смесь липидов также активи-
ровала кальцийзависимую протеинкиназу [Roberts, 1992].
Перекись водорода активировала [Stennis et al., 1998;
Tertyshnikova, Fein,
1998], а цГМФ ингибировала [Ванин,
1998] фосфолипазу С. Как полиеновые жирные кислоты,
так и их гидропероксиформы ингибировали кальциевые ка-
налы [Ninnemann, Maier, 1996].
Для липоксигеназной системы характерен один из наи-
более сложных механизмов регуляции (рис. 47). На фосфо-
липазу А
2
могут действовать МАР-киназы, так как они спо-
собны вызывать фосфорилирование и вследствие этого ак-
тивацию этого ключевого фермента [Hunter, 1995]. Повы-
шение концентраций фосфатидной кислоты [Ryu,Wang,
1998], а также ионов Са
2+
[Scherer, 1996a,b]
активировало
Рис. 45. Регуляция функционирования фосфатидатной сигналь-
ной системы интермедиатами других сигнальных систем
ЛФ - лизофосфатиды; МФЛ - мембранные фосфолипиды; ПКС -
протеинкиназа С; ПЛ - плазмалемма. Остальные обозначения - см.
рис. 42