ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 12.12.2023
Просмотров: 241
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
ААФ брадикининнің ыдырауын туғызатын кининаза II ферментіне ұқсас (1сызба) Брадикинин – микроайналу мен иондарды тасымалдауды реттеуге қатысатын күшті вазодилататор. Брадикининнің өмірсүру уақыты өте қысқа және қанағысында (тіндерде) өте аз концентрацияда болады; сондықтан ол жергілікті гормон (паракриндік реттеу) сияқты әсер көрсетеді. Брадикинин жасушаішілік Са2+ дің көбеюін тудырады , NO-синтетазаның кофакторы болады, ол эндотелийді босаңсытатын факторды түзуге қатысады (азот оксиді немесе NO). Эндотелий босаңсыту факторы тамырлардың жиырылуын тежейтін және тромбоциттерді біріктіретін қызметімен қатар митоздың ингибиторы және тамырлардың тегіс бұлшық еттерінің пролиферациясын туғызады да, антиатерогендік әсермен қамтамасыз етеді. Сонымен қатар брадикинин ПГЕ2 және ПГI2 (простациклин) тамыр эндотелиясының синтезін белсендіреді- олар қуатты вазодилаторлар және тромбоцитарлы антиагреганттар болып табылады.
Сонымен, брадикинин және кинин жүйесі РААЖ ға қарсы әсер етушілер болып табылады. ААФ ты тежеу жүрек тіндері мен тамыр қабырғаларында кининдердің деңгейін жоғарылатады, ал ол антипролифератты, антиишемиялық, антиатерогенді және антиагрегантты әсерді қамтамасыз етеді. Кининдер қан ағымының жоғарылауына көмектеседі, шумақты сүзгілеудің жылдамдығын өзгертпей ақ диурез бен натрийурезді қамтамасыз етеді. ПГЕ2 мен ПГI2 де диуретикалық және натрийуретикалық әсер көрсетеді және бүйректік қан ағысын жоғарылатады.
Іштегі нәрестенің сыртқы тынысалу жүйесі және қауіпті факторлар.
Іште жатқан нәрестенің дамуы кезінде тынысалу жүйесінің құрылымдық - қызметтік ерекшеліктері гетерохромдық(әртүстілік) түрде қалыптасады. Нәрестенің туылар алдындағы тынысалу жүйесі оның жаңа сыртқы ортаға сәйкес келетін метаболикалық керекті алмасу процесін қамтамасыз етуге дайын болуы керек.
Сыртқы тынысалу ағзалары: көмей, трахея(кеңірдек), бронх(кеңірдек тарамы) және өкпе - ерте эмбрионалды кезеңнен дами бастайды. 3-4- апталық нәресте өмірінде олар ілгері шыққан(дивертикула) эктодермалды өңеш түтігі сияқты болады. Осы ілгері шыққан каудалды аяғында томпақ кеңістік пайда болады , ол 4-ші аптада оң және сол жаққа бөлінеді (келешек оң және солжақ өкпе). Әр бөлік кішкентай тарауларға бөлінеді (өкпенің келешек бөліктері). Өкпе тіндерінің дивертикулаларының дамуы кезінде олар аяқ жақтарында шар тәрізді кеңістіктерді түзеді(бронхтың әртүрлі тарамдарының бастамасы ) және қоршаған мезенхемаға біріге бастайды. Мезодермалды мезенхималардан тегіс бұлшық етті талшықтар және бронх шеміршектері түзіледі. 6-шы аптада бойлатылған бөліктер түзіле бастайды, 8-10-шы апталарда – сегменттерге бөлінген бронхтар, ал 16-шы аптада - респираторлық бронхиоллалар түзіледі. Нәрестенің дамуының10-шы аптасында трахеяның және бронхтың шеміршекті каркасы түзіледі. 10-13-ші аптада тынысалу жолының шырышты бездері, кірпікті эпителийлері, бокал тәрізді жасушалары дами бастайды. Түптеп алғанда аталған процестер өкпенің бездік сатыларының дамуына сәйкес келеді.
Бронхтағы қуыс (қайтаканалдану сатысы) нәресте дамуының 16-шы аптасынан кейін түзіледі. Осы кезеңде респираторлық бронхиоллалар мен альвеолярлы жолдар қалыптаса бастайды, ал 24-ші аптада келешек ацинустардың қалыптасуы басталады (саккулярлы кезең, немесе альвеолярлы саты ). Сондықтан, 22- 26- апталарында іштегі нәрестенің өкпесі газалмасуына қабілетті болады. Бірақ альвеоланың нақты қалыптасу уақыты нәресте туылғаннан кейін басталады және бірнеше жасқа дейін созылады (8-жасқа дейін). Лимфатикалық тамырлар 9-10-шы апталарда дами бастайды, әуелі өкпенің түбірінен басталады. Кейінірек (13-ші аптадан) бронхтың шырышты қабатты бездері дами бастайды. Өкпе қантамырлары нәрестенің тіршілігінің 20-шы аптасында дами бастайды. Өкпенің васкуляризациясы(тамырлануы) 26-28-ші апталарда қарқынды жүреді, бірақ тамырлық жүйе толық нәресте туылар алдында қалыптасады.
Альвеолалардың сырты біртіндеп нәруыз-липидтік қабатпен, немесе беттік активті зат – сурфактантпен жабылады, оның рөлі альвеолалардың қабырғасын және ауа жолдарын және ауаны жабатын сұйықтықпен аралықта беттік керілу күшін төмендету. Сурфактант – ол нәруыз бен майдың қоспасы. Майлар негізінен лецитиннің туындылары(негізінен – дипальми-тоилфосфатидилхолин), олар бір- бірінен тебіседі де, альвеолаларда беттік керілу тудырады (бір- бірін жабыстырмайды). Сурфактанттың 40% фосфолипид, 5 % ға жуығы нәруыз. Сурфактанттың нәруыздары альвеоло-циттер арқылы секрецияланады.
Сурфактанттың жетіспеушілігі генетикалық патологиядан тууы да мүмкін, өкпеге ауа түскеннен кейін альвеолалар бүрісіп қалады, бұл тынысалудың ауыр бұзылу түріне әкеледі, емдемесе жаңа туған нәрестенің өліміне әкеледі. Сурфактанттың өкпедегі қызметі 14-ші аптада біліне бастайды. Бірақ, сурфактанттың қабаты 0,1-0,3 мкм қалыңдықтай жұқа болады және ол альвеолаларды бала туғаннан кейін ғана жаба алады, ол альвеолоцит II түрінде нәрестенің іште дамуының соңғы кезеңінде синтезделе бастайды. Сурфактанттың синтезіне метил- және фосфохолинтрансфераза қатысады. Метилтрансферазаны өкпеде іштегі нәресте дамуының 22-24-апталарында байқауға болады, бұл ферменттің белсенділігі нәресте туылар кезінде арта бастайды. Фосфохолинтрансферазаның өкпедегі белсенділігі жүктіліктің 35-аптасында байқалады. Сурфактанттың секрециясын арттыруға қанағымында айналып жүрген глюкокортикоидтар , катехоламиндер, простагландинЕ2 әсер етеді. Мысалы, егер әйел жүктілік кезінде глюкокортикоидтарды қабылдаса, нәрестедегі сурфактант өкпеде өте ерте пайда болады. Жүкті әйелдердің стресс жағдайында (босануы, ауыру және т.б. ) сурфактанттың өкпеде дамуы артады, ол ана ағзасындағы глюкокортикоидтардың белсенділігінің артуымен байланысты болуы мүмкін. Сурфактанттың арқасында жаңа туған нәрестенің алғашқыда өкпесінің тынысалу бұлшық еттерінің жазылуына күшсалу аз жұмсалады, ол ары қарай тіршілік өмірінде жалғаса береді. Сонымен қатар, сурфактант тынысалу кезінде өкпе көлемінің айтарлықтай өзгеруіне қарамастан, альвеолалар жұмысының тұрақылығын қамтамасыз етеді.
Іште жатқан нәрестеде сыртқы тынысалу жұмыс істемейді. Іштегі нәрестенің жоғарғы тынысалу жолы амниотикалық сұйықтықпен толтырылған. Альвеолалар мен бронх жазылған түрде болады, себебі олар өкпелік сұйықтықтармен толтырылған және кезекте басымырақ альвеолоцит II түрінде секреттеледі және жетілуші өкпенің ішкі ортасы болады. Амниотикалық және өкпе сұйықтықтары араласпайды, себебі бұған жіңішке өңеш кедергі келтіреді.
Жатырдың шырышты қабатында аналық жасушаны ұрықтандырғаннан кейін және оны имплантациялау уақытына дейін аналық жасуша оттекті диффузия процесі кезінде 02 пен СО2 жатырдың шырышты қабатынан алады. Ұрықтың плацентарлы даму кезінде О2 тің тасымалдануы басқа механизммен іске асады. Жүкті әйелдердің артериялды қанындағы О2 тің қысымы 110 мм сын.бағ. 14,6 кПа, және бұл қан жатырастылық артериямен плаценттің түкті кеңістігінің аралығымен келеді. Плацентке басқа жағынан кіндік артериясы арқылы аралас қан ұрықтың ішек аортасынан келеді. Ондағы О2 қысымы плацентарлы ана қанының қысымына қарағанда төмен. Концентрациияларының айырымы нәтижесінде О2 хорионның түкті капиллярлары қанына диффузияланады. Плацентте О2 тің диффузиясы өкпеге қарағанда төмендеу болады (өкпе мембранасына қарағанда плацентарлық мембрананың қалыңдығы 5-10 есе қалыңдау). Ұрықтың артериалық қанында оттектің (О2 ) қысымы 20-50 мм сын. бағ. (3,3-6,7 кПа) (жекеше вариациялары болуы мүмкін). Бұл уақытта гемоглобин оттекпен орташа шамамен 65% (40 тан 80%ға дейін) қанығады. Осыған қарамастан, ұрық қанындағы гемоглобиннің мөлшері және оның оттектік сиымдылығы ересектердің шамаларына жақын , плаценттен кететін қандағы оттектің көлемі салыстырмалы төмен, яғни 80-140 мл/л дей болады.
Феталды гемоглобин диссоциациясының қисығы солға қарай ығысқан – яғни О2 тің қысым мөлшері төмен аймағына қарай. Сонымен қатар, ересек адамдарға қарағанда, ұрық оксигемоглобині диссоциациясының қисығы өте тік болып келеді. Оксигемоглобин диссоциациясы қисығының солға қарай ығысуы феталды гемоглобиннің оттекке деген өте жоғары ынтықтығын көрсетеді. Бұл негізінен ұрық эритроциттеріндегі 2,3-дифосфоглицераттың аз мөлшері болуымен байланысты. 2,3-ДФГмен феталды гемоглобиннің байланысу қабілеті ересек адамдардағы байланысудың 40% ын құрайды. Ал ересектерде, керісінше, бұл зат гемоглобиннің О2 ке ынтықтығын өте коп дәрежеде төмендетеді. Мұндай ұрық оксигемоглобинінің диссоциациялану қисығының сипаты тіндерді оттекпен жақсы қамтамасыз ету дәрежесін көрсетеді.
Ұрықтың артериалды қанындағы СО2 нің қысымы 38-45 мм сын. бағ. (5-6 кПа) [ кейде – 32-33 мм сын. бағ. (4,3-4,4 кПа)], яғни ұрықтық қан нормокапниямен (қалыпты көмірқышқыл газымен қанығады) сипатталады. Жүкті әйелдерде гипокапния (гипервентиляцияның нәтижесінде) байқалатын болғандықтан, гематоплацентарлық кедергіде СО2 нің айқын концентра -циялық градиенті орын алады, ол ұрық қанынан оның қарқынды кетуіне мүмкіндік береді.
СО2, ұрық қанында ересектердікі сияқты үш жолмен тасымалданады: еріген күйде, бикарбонат және карбаминді қосылыстар (гемоглобиннің амин топтарымен байланысқан) түрінде. Бірақ фетальды гемоглобин ересектердің гемоглобинімен салыстырғанда қарқынды түрде карбаматты түзеді, ол көмірқышқыл газының ағзадан шығыуына мүмкіндік береді. Ұрықтың аралас қанында СО2 нің мөлшері 400-500 мл/л аралығында болады.
Ұрықтың эритроциттеріндегі карбоангидразаның мөлшері онша көп емес, сондықтан оның СО2 ні тасымалдаудағы белсенділігі, ұрықтың биологиялық мембранасынан өтуі, ересектердің 20-25% ын құрайды. Ферменттің белсенділігі жүктіліктің соңғы кезеңінде арта бастайды және ересектердің дейгейінің 30-50% ын құрайды. Карбоангидразаның мұндай белсенділігі, ұрықтағы СО2 нің газ алмасуының физиологиялық қалыпты жағдайы үшін, көмір қышқылының түзілуі мен диссоциациясына жеткілікті (СО2 + Н2О → Н2СО3 → H+ + НСО3-). Осының нәтижесінде ұрықтың аралас қанының рН- 7,13-7,23, артериалдық - 7,24 -7,32 (ересектердікіне қарағанда төмен) тең болады. Антенаталды кезеңде анаэробты процесстер қарқынды жүреді, ол ұрықтың аралас қанында сутек иондарының жоғары концентрация -сын тудырады, себебі оған зат алмасу кезінде қышқылдық өнімдер көп түседі (метаболикалық ацидоз), соның ішінде сүт қышқылы. Сондықтан ұрық қанында сілтілік резервтің шамасы төмен болады.
Сонымен, ұрықта О2 пен СО2 нің қанда тасымалдануы, ересектерге қарағанда, келесі ерекшеліктерімен: 1) О2 төменгі мөлшері және гемогло- биннің оттекпен қанығуының жеткіліксіздігімен; 2) СО2 қысымының ересектерге жақындығымен немесе аздап төмендігімен; 3) қанда сутек иондарының жоғары концентрациясымен; 4) карбоангидразаның белсенділігінің төмендігімен; 5) сілтілік резерв деңгейінің төмендігімен сипатталады. Ағзадағы ұрықта осындай ерекшеліктер газалмасу гомеостазын сақтай алады.
Өкпенің атқаратын қызметтері
Өкпенің тынысалу қызметіндегі ең маңыздысы атмосфералық ауа мен қан арасында газалмасуымен байланысты және ағзаның ішкі ортасы құрамының тұрақтылығын сақтау. Сонымен қатар, өкпе ауамен түскен ағзаға жат заттарды – ксенобиотиктерді залалсыздандырады, жасушалық иммунитетті сақтау реакцияларына қатысады және қышқылдық-негіздік күйдің тұрақтылығын сақтауға қатысады. Өкпеде өзінің қызметін атқаратын заттар синтезделеді (сурфактант жүйесінің компоненттері және биологиялық активті заттар), олар әртүрлі ағзалар мен тін жасушаларының осы заттарға сезімталдығын реттейтін заттар. Өкпе қаннан вазоактивті гормондарды алып, оларды бұзады немесе өзгертеді. Көрсетілген қызметтерді респираторлық емес өкпенің қызметтері деп атайды. Бұл тарау өкпенің респираторлық емес қызметтері және оның ерекшеліктеріне арналады. Өкпедегі газдардың тасымалдану процесстері «Тынысалудың биофизикалық негіздері»тарауында кеңінен қаралған.
Өкпеде зат алмасуының ерекшеліктері
Кез келген органдарда, тіндерде зат алмасу процесстері жүреді. Кәзіргі кезде негізгі метаболизм жолдары жеткілікті анықталған. Әртүрлі органдарда және тіндерде зат алмасуы жалпы бірдей және арнайылықты жүретін ерекшеліктері болады. Заттардың ыдырау процесстері (катаболизм) органикалық молекулалардан энергияны бөліп және оны макроэрг - зат ретінде немесе иондардың градиенті ретінде жасуша мембраналарында қорға жинау. Катаболизм процесі жүретін химиялық реакциялардың түрлеріне қарай энергетикалық өзгеріс экзергондік процеске жатады. Экзергондік процестерде энергия бөлінеді. Синтез процесі (анаболизм) кезінде жасушаларға және жалпы ағзаға керекті заттар түзіледі. Күрделі заттардың жай заттардан синтезделуі кезінде энергия жұтыла жүреді. Мұндай процесстер эндергондік деп аталады.. Анаболикалық реакциялар жасушалардың құрылыстары мен қызметтерінің белсенділігін қамтамасыз ететін заттарды түзеді. Кейбір жасушаларда синтезделген заттар сыртқа шығады және өзінің рөлін түзілген жерінен басқа алыс жерде атқарады.
Өкпе метаболизмінің ерекшеліктері туралы мәліметтер онша көп емес. Мұның себебі метаболизмде өкпе рөлі оның құрылысының ерекшеліктерімен байланыстылығы. Өкпе кеуде жасушаларының көп бөлігін алады, сонымен қатар өкпе массасы бүкіл ағзаның тек 1% ын ғана құрайды. Өкпе – қан толық бір айналым жасап, одан толық көлемімен өтетің жалғыз орган. Өкпенің 30 % дай массасы ондағы қанның молшеріне сәйкес келеді. Сондықтан, өкпенің массасы одан өтетін қанның көлеміне қарағанда кіші. Демек, қанға келіп түскен метаболиттердің концентрациялары мен одан шыққан метаболиттер концентрацияларының айырымы(артериялық -көктамырлы айырым) өте аз болғандықтан, бұл ағзада биохимиялық процесстер туралы мәліметтер мардымсыз. Бұл ерекшелік қан құрамындағы газ құрамының айырымында, бірақ бұл өкпенің метаболизмін көрсетпейді, тек оның газалмасу рөлін көрсетеді. Өкпе метаболизмін зерттегенде тәжірибелік модель жасауға тура келді ( in vitro). Мұндай моделдің мысалы ретінде оқшауланған перфузделген өкпенің моделі ұсынылды ( 1 сурет).
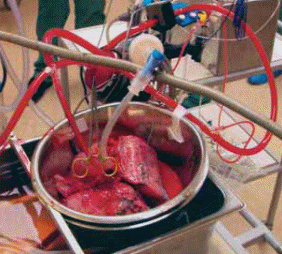
1сурет. Оқшауланған өкпенің моделі. Бір түтік өкпе артериясымен , ал екінші түтік өкпе көктамырымен байланыстырылады. Перфузделген сұйықтық өкпе арқылы айналып жүреді.
Ағзаның перфузиясы, жасанды ортада, каннан толық жуылуды қамтамасыз етеді. Сондықтан, өкпеге келетін және одан кететін сұйықтықтардағы заттардың мөлшерінің айырымы осы заттардың өкпе жасушаларындағы биохимиялық өзгерістерімен байланысты. Өкпе тіндеріндегі метаболизм процесстерін зерттейтін басқа модель өкпе тінін кесу, өкпе жасушаларының дақылдары(культуралары) мен органеллалардың субжасушалық бөліктерін ажырату. II типтегі альвеолоциттердің оқшауланған дақылдарын қолдану сурфактант өнімі үшін бұл жасушалардың рөлін сипаттауға мүмкіндік берді.
Өкпенің оттекті пайдалануы
Әртүрлі органдардың оттекті пайдалану жылдамдығын біле отыра, осы органда жүретін зат алмасуының қарқындылығы туралы пікір айтуға болады. Метаболикалық белсенді органдар оттекті көп мөлшерде , ал белсенділігі жоқ органдар оттекті аз мөлшерде пайдаланады. Өкпеден қанға көп мөлшерде оттек түседі. Бірақ көп уақытқа дейін өкпенің өзі қандай көлемдегі қанды пайдаланатындығы белгісіз болып келді. Бұған жауап өкпенің тіндерін перфузиялық кесінді жасау тәжірибесі арқылы алынды. Оттекті сіңіру жылдамдығы тіндердің таза құрғақ салмағына есептегенде 30-150 мкл О2 мин-1 г-1 тең, ол жүрек, бүйрек, бас миы, қалқанша бездері сияқты метаболикалық активті органдарға қарағанда, өте аз шама. Бірақ,қаңқа бұлшық еттерінің тыныштық жағдайында және метаболикалық белсенділігі төмен органдарға қарағанда бұл шама әлдеқайда жоғары. Сонымен, өкпенің оттекті пайдалану жылдамдығын орташа шамаға жатқызуға болады. Бұан, бүкіл ағзадағы өкпенің массасы 1%ды құрайды және ол 1% ағзаға түскен оттекті пайдалады деген тұжырымға сәйкес келеді. Өкпенің тіндері біртекті емес, сондықтан ондағы әртүрлі жасушалардың оттекті пайдалану қабілеттері де әртүрлі болатындығын айта кеткен жөн. Мысалы альвеолоциттердің II – түріне оттекті өте көп пайдалану тән.
Өкпе отын ретінде қандай субстраттарды пайдаланады ?
Физиологиялық жағдайда, барлық ағзалардың жасушалары үшін негізгі энергетикалық отынның көзі глюкоза болады. Энергетикалық отын көзі ретінде тек глюкозаны пайдаланатын тіндерді облигативті пайдаланушылар деп атайды. Бұған мысалдар ретінде жүйке тіндерін, ішек эпителийлерін, бүйректің милы қабатын, эритроциттерді алуға болады. Көптеген тіндер энергия көзі ретінде глюкозадан басқа субстраттарды – бос май қышқыл- дарын, кетон денелерін, амин қыщқылдарын да пайдалана алады. Бұл субстраттарды пайдалану жасушаларға глюкоза жетіспей жатқан жағдайда көбейеді - аштықта, қарқынды физикалық жұмыс жасағанда, суық климаттық жағдайда, кейбір гармоналды бұзылыстарда (қантты диабет). Мұндай тіндер глюкозаны факультативті пайдаланушылар деп аталады.
Глюкозаны факультативті пайдаланушыларға өкпе де жатады, яғни қалыпты жағдайда оның негізгі энергетикалық субстраты глюкоза болады. Аэробиоз жағдайында өкпеге перфузия әдісі арқылы тәжірибе жасағанда, глюкозаны пайдалану тіннің құрғақ массасының 40мкмоль с-1г-1 ын құрайды. Анаэробты метаболизмге ауысқанда өкпенің глюкозаны жұту қабілеті 2 есе артады.
Барлық ағзаларға қарағанда өкпе ең көп оттекпен қамтамасыз етілетін ағза. Осыған байланысты өкпе тіндерінде анаэробты гликолиздің қарқынды жүруі күтпеген жағдай сияқты. 2 суретте өкпенің перфузиялық метаболизмі арқылы анықталған глюкозаның көміртек атомдарының «тағдыры» көрсетілген.
2 сурет. Оқшауланған өкпе өнімдерінің құрамына глюкозаның көміртек атомдарының енгізілуі
Метаболизмге қатысатын глюкозаның жартысына жуығы лактатқа айналады, ол гликолиздің маңызды рөлін және Кребс циклінің шектеулі рөлін растайды. Глюкозаның 22% ғана өкпеде СО2 мен H2O дейін аэробты ыдырауға қатысады. Осының 1/4 мөлшерінен цитоплазмада пентозды цикл арқылы көмір қышқылының бір бөлігі түзіледі және 3/4 бөлігі үш карбон қышқылы циклі арқылы митохондрияда изоцитраттың және альфа-кетоглутараттың тотығудан декарбоксилдену реакцияларынан түзіледі. Өкпеде гликолиздің қарқынды жүру мүмкіндігін, оны өкпе тіндері жасушаларының құрамына қарай өте гетерогенді болуынан және Кребс циклі реакциялары жүретін көптеген жасуша түрлерінде митахондриялар саны онша көп еместігінен деп түсіндіреді. Глюкозадағы көміртек атомдарының үлкен бөлігі нәруыздардың және нуклеин қышқылдарының құрамына кіреді (18%). Глюкозаның аздаған мөлшері май қышқылдарына (4%), басқа липидтерге (3%) және полисахарид- терге (гликогенге) (5%) айналады. Өкпе глюкозадан басқа май қышқылдарын да тотықтырады.
Өкпені энергиямен қамтамасыз етудегі аэробты метаболизмнің үлесі
Жоғарыда көрсетілген нәтижелерден, глюкозаның көп мөлшері лактат түзе, анаэробты ыдырауға ұшырайтындығын көрсетеді. Бұдан өкпені аэробты метаболизм бойынша энергиямен қамтамасыз етуге қосатын үлесі онша маңызды емес деген болжам шығады. Бірақ, өкпе перфузиясында аэробты метаболизмге игибиторлардың әсері, тынысалуды ажыратушылар мен митахондриялардағы фосфорлануда түзілген АТФ мөлшері мен АТФ/АДФ қатынасын қараудағы тәжірибе нәтижелері мұндай қорытынды жасай алмайды, керісінше өкпені энергиямен қамтамасыз етуде аэробты метаболизмнің маңызды рөлін көрсетеді.
Перфузиялық оқшауланған өкпеде АТФ мөлшері (тіннің құрғақ салмағы
10мкмоль/г) осы сияқты басқа да көп тіндерде, аэробты метаболизм түрінде жұмыс істегенде, АТФ/АДФ қатынасы 8,5 ке тең болады. Өкпеде тынысалу ингибиторы -СО әсерінен АТФ мөлшері мен АТФ/АДФ қатынасы 2-3 есе төмендейді. Өкпе тіндеріне тынысалу ажыратушыларымен әсер еткенде және динитрофенолмен фосфорлағанда да АТФ мөлшері мен АТФ/АДФ индексі төмендейді, бірақ СО әсеріне қарағанда олардың әсері азырақ болады.
Өкпедегі энергияға тәуелді процесстер
Өкпеде бұлшық еттің жиырылуы, ішек эпителийлерінен сіңірілу немесе бүйрек каналдырынан сіңірілу, жүйке импульсының өтуі сияқты энергияға тәуелді процестер жүрмейді. Өкпенің негізгі қызметі – энергияға тәуелсіз газдарды тасымалдау. «Тынысалудың биофизикалық негіздері» тарауында альвеолярлы ауа мен қан арасына тасымалданатын газдар жәй диффузиялық процесс деп айтылған. Бірақ кейбір өкпеде жүретін процесстер, мысалы, бронхқа цилиарлы тасымалда тегіс бұлшықет элементтерінің жиырылуы, сурфактантатың негізгі компанентінің синтезі- дипальмитоилфосфатидилхолиннің және оның альвеоалардың қуысынан секрециялануы сияқты процестер қарқынды түрде энергиямен қамтамасыз етуді керек етеді және гипоксия кезінде баяулайды.
Сурфактант жүйесі
Сурфактант – табиғаты липопротеидті, су-ауа беттік аралығында орналасқан, альвеолалардың ішкі бетінде төселген және дем шығарғанда алвеолалардың беттік керілуін төмендету арқылы олардың бүрісуіне кедергі жасайды. Сурфактант өкпенің арнайылықты әртүрлі жасушаларынан – екінші дәрежедегі альвеолоциттермен секреттелінеді. Туа біткен сурфактанттың жетіспеушілігінен жаңа туылған нәрестелерде өкпенің ашылмауынан - респираторлық дистресс-синдромы (РДС) байқалады. Сурфактанттың негізгі компоненті фосфолипидтер, нәруыздар және полисахаридтер. 1 кестеде сурфактанттың химиялық құрамы туралы мәліметтер берілген.
Липидтер
Липидтер сурфактант массасының 90%дан көбін құрайды және оның жартысы қаныққан глицерофосфолипид – дипальмитоилфосфатидил- холиннің үлесіне тиеді. Сурфактантатың екінші компоненті басқа глицеро- фосфолипид – фосфатидилглицерин болып табылады, оның құрамына қанықпаған май қышқылдарының қалдықтары кіреді. Сурфактанттың фосфолипидтері альвеолалардың ішкі қабатын түзеді. Фосфолипидтердің полярлы топтары сулы ерітінділерге , ал гидрофобты май қышқылды құй- рықтары ауаға қарай бағытталып, су- ауа аралығындағы беттік керілуді төмендетеді.
1кесте. Сурфактанттың негізгі компоненттері
| Фосфолипидтер: 85% | Жеке фосфолипидтердің (%) және нәруыздардың шартты бірлікте берілген мөлшері |
| 47,0 |
| 29,3 |
| 11,6 |
| 3,9 |
| 3,3 |
| 1,5 |
| 3,4 |
| Бейтарап липидтер: 5% | |
| Холестерол, бос май қышқылдары | |
| нәруыздар: 10% | |
| ++++ |
| + |
| + |
| ++ |
| |
Сурфактант құрамына аздаған мөлшерде бейтарап липидтер мен холестерин кіреді. Сурфактанттың барлық липидтері II типтегі альвеолоциттермен син- тезделеді, олар ламеларлы дене деп аталатын секреторлы органеллаларда жинақталады.
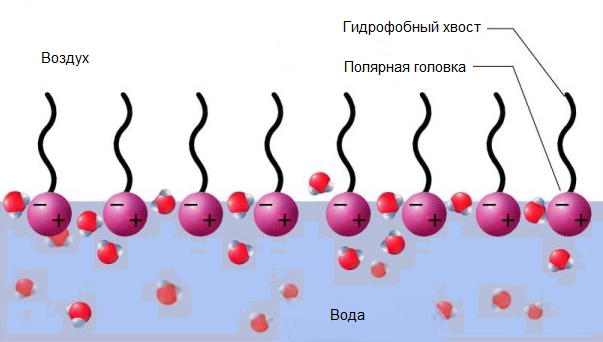
3 сурет. Су – ауа беттік аралығында альвеолалардың ішкі қабатында түзілген фосфолипидтердің беттік моноқабаты
Нәруыздар
Сурфактант массасының 10% нәруыздардың ұлесіне келеді. Бұның жартысы қан плазмасының нәруыздары, екінші жартысы сурфактант нәруыздары – аполипопротеиндер А (SP-A), В (SP-B), C (SP-C) және Д (SP-D). Ағылшынша аббревиатура SР -surfactant protein- яғни сурфактант нәруыздары дегенді көрсетеді.
-
SP-A және SP-D бактериялардың беттеріндегі көмірсу домендерін танып және байланыстыру қабілетіне қарай туа біткен иммунитет реакция- ларына қатысады. Байланыстыру микроорганизмдердің макрофаг- тарының фагоцитозымен қоса жүреді. Сонымен қатар SP-A сурфактант синтезін реттейтін теріс кері байланыс механизміне де қатысады. -
SP-B және SP-C - гидрофобты мембраналық нәруыздар, альвеолалардың бетінде сурфактанттардың дұрыс орналасуына көмектеседі және өкпеде биофизикалық механизмдердің қызметін қамтамасыз етеді. Бұл нәруыздардың туа біткен жетіспеушіліктері өкпе жұмысының ауыр бұзылыстарымен қоса жүреді: SP-B жетіспеушілігі өкпе жетіспеу -шілігімен қоса жүреді, ал SP-C тің жетіспеушілігі интерстециальды пневмонияның (қабынудың)дамуына мүмкіндік туғызады. -
Сурфактанттың аполипопротеиндері альвеолоциттердің IIтүрінің көмегімен түзіледі.Трансляциядан кейінгі өзгерту нәтижесінде бұл нәруыздар үш өлшемді құрылымға айналады (4 сурет) және жасушада олар ламелярлы дене түрінде оралады– диаметрі 1 мкм табиғаты липопротеидтер болатын концентрленген сақина тәрізді.
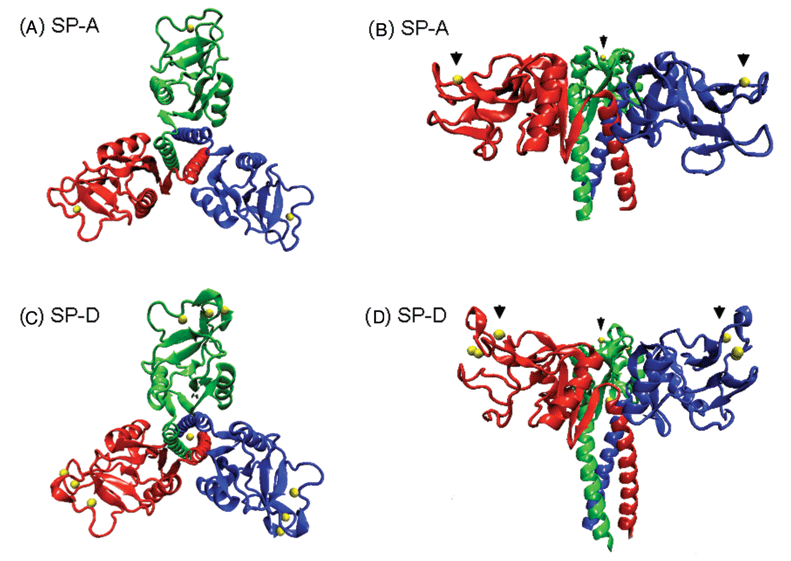
5 сурет. Сурфактант аполипопротеиндерінің үш өлшемді моделі
Сурфактанттың атқаратын қызметтері
-
Өкпенің созылмалығы ұлғайады; -
Ателактазадан(өкпе тіндерінің бүрісуі) сақтандырады -
Иммундық қорғанысқа қатысады .
Альвеолаларды судағы ауа көпіршіктері ретінде қарастыруға болады. Су- ауа беттік аралығында түзілген беттік керілу күші альвеоларды қысуға тырысады. Сондықтан оның көлемінің кішіреюінен сақтандыру үшін альвеолаларға белгілі бір ауа қысымы Р керек, оны Лаплас теңдеуімен есептеуге болады. Теңдеу қысымның (Р) беттік керілу күшіне(γ) және альвеолалардың радиусына (r)тәуелділігін көрсетеді: