ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 12.12.2023
Просмотров: 242
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
Өкпенің созылмалылық қасиеті
Өкпе тіндерінің созылмалылығын оның бүкіл кеуде жасушаларын алатындық қабілеті бар деп қарауға болады. Өкпенің негізгі созылмалылық рөлін сурфактант атқарады. Соңғы дем шығару фазасында сурфактант қабатының жиырылуы альвеолалардың бетіндегі беттік керілудің төмендеуіне әкеледі, ол нөлдік деңгейге дейін жетуі мүмкін. Дем алған уақытта өкпенің ашылуы(кеңеюі) жақсарады. Сурфактант беттік керілу күшін төмендетуі арқасында, ол сұйықтықтың альвеолаларда жиналуына кедергі жасайды.
Альвеолалар мөлшерінің реттелуі
Дем алуда альвеолалар көлемі үлкейеді. Бұл сурфактант қабатының созылуымен және беттік керілу күшінің жоғарылауымен қоса жүреді , ол альвеолалардың ары қарй үлкеюіне кедергі жасайды. Сурфактанттың беттік керілу күшінің жоғарылауы альвеолалар арасындағы қуыстардың үлкеюіне де әкеледі. Егер кейбір альвеолалардың мөлшері өте көп үлкейіп, ал басқалары сол қалпында қалса , онда осы альвеолалардағы беттік керілудің жоғарылауы олардың ары қарай үлкеюіне кедергі жасайды. Осының нәтижесінде дем алғанда альвеолалардың арасындағы қуыстар бірдей үлкейеді. Осындай заңдылық дем шығарғанда да байқалады және дем шығарғанда – егер кейбір альвеолалар көп мөлшерде жиырылса, онда беттік керілудің төмендеуі олардың ашылуына көмектеседі.
Альвеолалардың қуыстарына сұйықтықтың түсуінен сақтандыру
Беттік керілу күші капиллярлардан сұйықтықтардың альвеолалар қуысына шығуын жоғарылатады, бірақ сурфактант қабаты бұл күшті төмендету арқылы сұйықтықтың капиллярлардан шығуынан сақтандырады.
Иммунндық қорғауға қатысуы
Өкпе өте осал орган. Ағзаның ішкі ортасымен сыртқы қоршаған ортаның шегінде орналасқандықтан, өкпе ауамен байланысу үшін өте үлкен бетке ие болады, сондықтан қан мол келеді және қан тамырлары өткізілген эпителиидің жұқа қабаты болады. Тәулігіне өкпеден 11000 литр атмосфералық ауа өтеді. Өкпе арқылы ішкі ортаға қандай мөлшерде микроорганизмдер, патогендер, аллергендер және ауаны ластайтын заттар түсетінін көзге елестету оншалықты қиыншылық тудырмайды. Бірақ бағына қарай өкпе өзін қорғайтын күшті механизмдермен жабдықталған. Олар микроорганизмдерді және қатерлі заттарды шығарады және зарарсыздандырады. Солардың ішіндегі механизмдердің бірі, туа біткен және жолай пайда болған иммунитет реакциялары маңызды рөл атқарады. Мұндай қорғаныштық механизмдердің ішінде маңызды рөл сурфактант нәруыздарына беріледі. Сурфактант аполипопротеиндері SP-A мен SP-D коллектин тұқымдастығына жатады, олар бауырда синтезделетін лектиндерді, маннозаның қалдығын тануға қабілетті заттардың құрамына кіреді. Коллектиндердің N-тізбегі соңында коллагеннің құрылысына ұқсас аймақтары болады, ал С-соңында бактериялардың бетінде көмірсуларды байланыстыратын домен болады (5сурет).
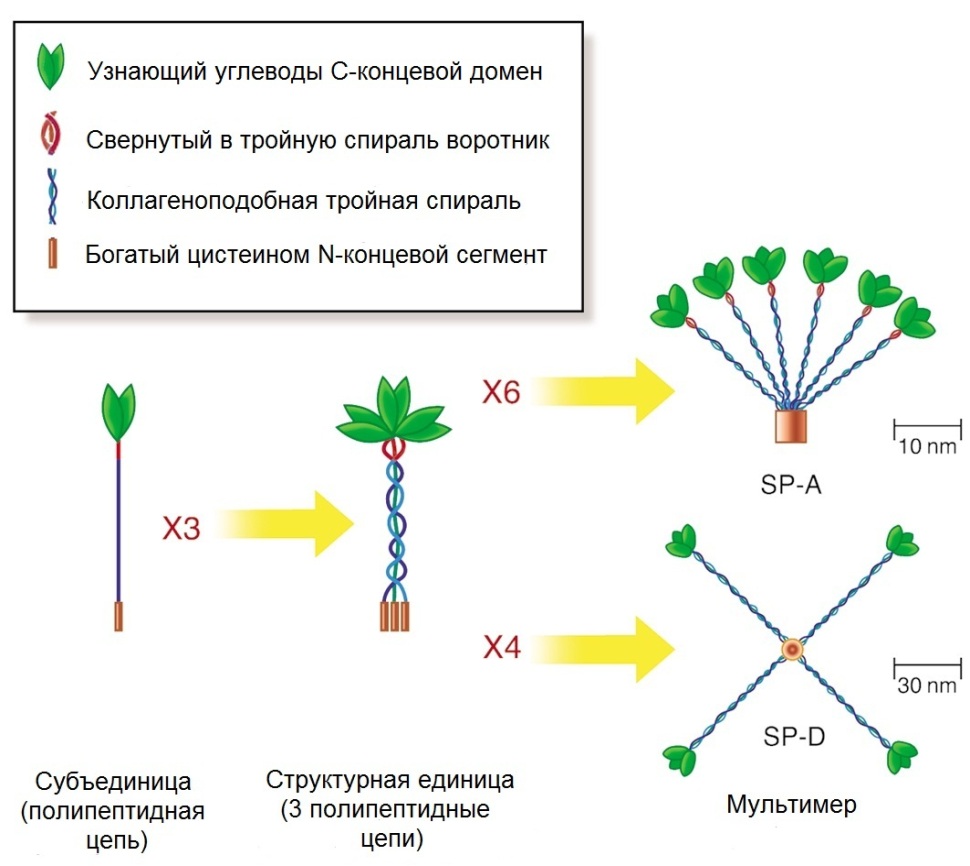
5сурет. Коллектиндер тұқымдастарының құрылысы. SP-A 6 құрылымдық бірліктерден тұрады (Х6), олар «гүл шоғына» ұқсас құрастырылған. SP-D төрт өлшемді құрылымды (Х4) .
Бактериялардың бетіндегі көмірсулар детерминанттарын тани білу арқылы, SP-A мен SP-D микроорганизмдерді бірден өлтіреді немесе макрофагтар мен моноциттердің фагоцитозы арқылы іске асырады(6 сурет). Сонымен қатар SP-A мен SP-D цитокиндердің қабыну өнімдерін де реттей алады.
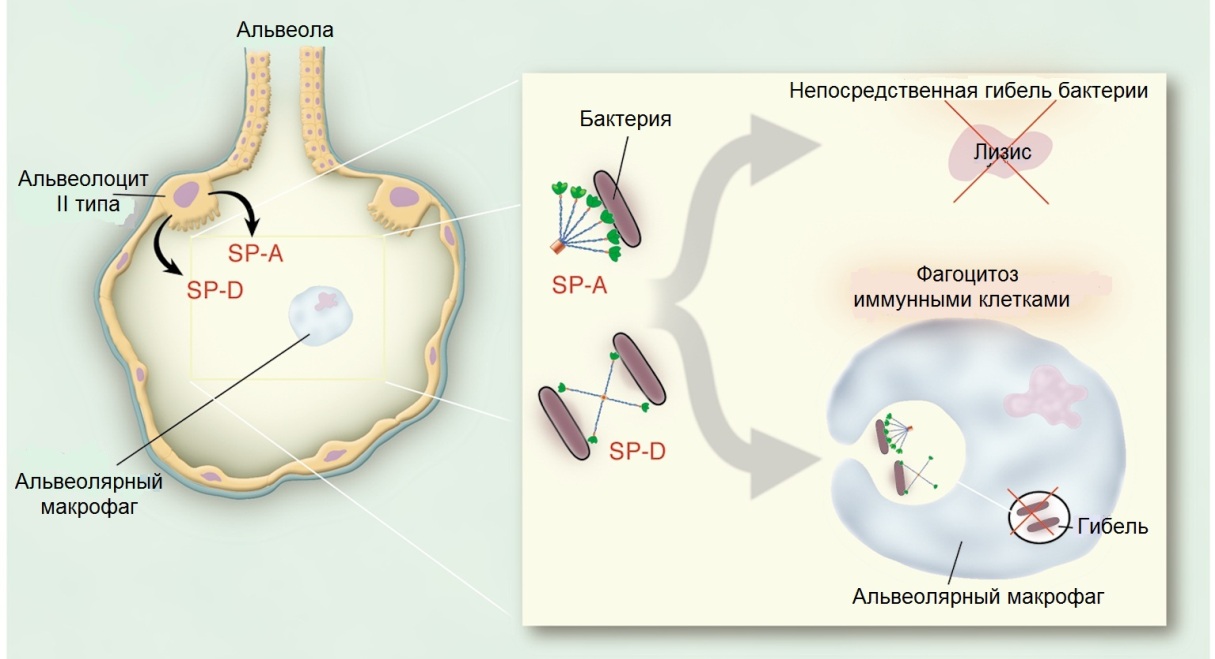
6 сурет. Альвеолоцит II түрінен SP-A мен SP-D ның синтезі және олардың иммундық қорғаудағы рөлдері
Ксенобиотиктердің метаболизмі
Ағзаға тыныс алу кезінде ауамен бірге көп мөлшерде улы заттар түседі. Мұндай заттар тіршіллікке бөтен заттар, сондықтан олар ксенобиотиктер деген атқа ие болды. Ксенобиотиктер өндіріс жұмыстарының нәтижесінен түзіледі және қоршаған ортаны ластайды. Көптеген ксенобиотиктер химиялық табиғаты жағынан ароматты көмірсуларына жатады немесе олардың туындылары болып табылады. Адам ағзасына өкпе арқылы түсетін арнайылықты ксенобиотиктерге шылым түтінінің компоненттері, мысалы бензпирен, жеңіл автомобилдердің пайдаланылған газдары, қазан мұржасынан шыққан булар(газдар) және т.б. жатады. Физико-химиялық қасиеттері жағынан мұндай ксенобиотиктер – гидрофобты қосылыстар. Олар ағзаға түскенде жасушалардың гидрофобты фазасында, яғни биологиялық мембраналарында жинақталады, сондықтан олар – канцерогенді, мутагенді, аллергеногенді, тератогенді және т.б. ағзаға жағымсыз әсер етеді. Ксенобиотиктер суда нашар еритін болғандықтан ағзадан сұйықтықтармен бірге шығып кете алмайды, яғни кумулятивті ( концентрациясы жинақталған) эффект көрсетеді. Ксенобиотиктерді ағзадан шығару үшін олардың молекулаларының полярлылығын арттыру керек. Ол үшін ксенобиотиктерді тотықтыру керек және тотыққан кезде түзілген полярлы оттегісі бар топқа гидрофильді молекулаларды – адъюванттарды байланыстыру керек. Бауырдың залалсыздандыру қызметі екі реакцияның үйлесіміне –тотығу мен конъюгацияға негізделген.
Ксенобиотиктердің тотығуын (залалсыздандырудың бірінші фазасы) мембрананың эндоплазматикалық ретикулумының монооксигеназды жүйесі іске асырады
. Монооксигеназды жүйенің негізгі компоненті гемопротеид – цитохром Р-450 болады. Бұл нәруыз молекулалық оттекті белсендіреді және оттектің бір атомын ксенобиотик молекуласының құрамына енгізеді. Мұндай реакцияларды гидроксилдену реакциялары деп атайды, себебі гидрофобты молекуланың орнына гидроксил тобы бар полярлы молекула пайда болады (7 сурет).
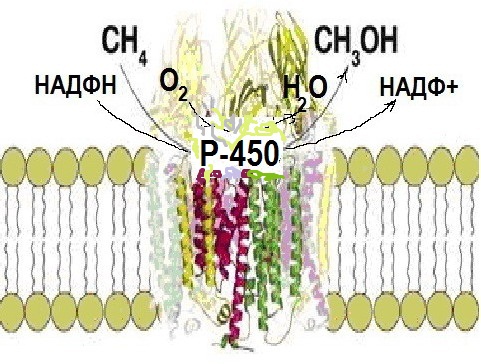
7 сурет. Монооксигеназды жүйенің жұмыс істеу принципі .
Цитохром Р-450 молекулалық оттекті белсендіреді және бір атом оттекті гидрофобты молекуланың құрамына ендіреді.
Оттектің екінші атомы сутекті тотықтыруға жұмсалып, судың молекуласы түзіледі.
Залалсыздандырудың екінші фазасында трансфераза ферменттері тотыққан ксенобиотикті адъюванттың(иммундық жүйені белсендіруші) гидрофильді молекуласымен конюгациялау реакциясын жүргізеді. Адьювант рөлдерін глюкурон және күкірт қышқылдары, амин қышқылдары және де басқа заттар атқарады.
Әртүрлі ксенобиотиктер үшін цитохром P-450дің арнайылықты түрлері болады, ксенобиотик ағзаға түскенде сәйкес геннің экспрессиясы жүреді және осы ксенобиотик түріне арнайылықты цитохрома Р-450 синтезделеді. Супертұқымды Р-450 цитохромдары бірнеше тұқымдарды біріктіреді, олар бірнеше ондаған гендермен кодталады.
Ең қуаттыы монооксигеназды жүйе бауыр жасушаларында болады. Өкпеде де монооксигеназды жүйе болады, бірақ оның қуаттылығы бауыр монооксигеназды жүйесінің 25% ғана бола алады. Сонымен, ксено- биотиктерді ағзада залалсыздандыруда өкпе де елеулі үлес қосады.
Биологиялық белсенді заттардың (гормондар және гормонға жатпайтын биореттеушілер) және дәрілік заттардың метаболизмдері
Өкпе биологиялық белсенді заттар (БАЗ) мен дәрілік заттардың метаболизміне екі механизм арқылы белсене қатыса алады:
-
Қанда айналып жүрген БАЗды және дәрілік препораттарды ұстап алып оларды химиялық өзгерістерге ұшыратады, сонымен қоса олардың биологиялық белсенділіктерін өзгертеді; -
Бірқатар БАЗ ды синтездейді.
Өкпенің БАЗ бен дәрілер метаболизміндегі рөлі 2- кестеде жинақталған. Бірқатар БАЗ бен дәрілік заттар өкпеде белсенділіктерін жоғалтатыны анықталған. Керісінше кейбір БАЗдар өкпеде метаболикалық түрөзгеріс-тердің нәтижесінде белсендіріледі. Өкпе негізі липидтік биореттеушілерді – простагландиндерді синтездейді. Кейбір БАЗдың биологиялық белсенді– ліктері өкпеден өткеннен кейін өзгермейді.
2 кесте. Өкпенің бірқатар БАЗ метаболизміне қатысуы
| Өкпемен активсізденетін заттар | Гормондар және гормонға жатпайтын БАЗ: |
| Серотонин | |
| Ацетилхолин | |
| Норадреналин | |
| Брадикинин | |
| Простаглангдиндер: Е1, Е2 , Е2ά | |
| Дәрілік препараттар: | |
| Аминазин | |
| Имипрамин | |
| Нортриптилин | |
| Пропранол | |
| Сульфаниламидтер | |
| Циклизин және хлорциклизин | |
| Димедрол | |
| Галюционгендер: | |
| Лизергин қышқылының диэтиламиді | |
| Өкпемен белсендірілетін немесе синтезделетін заттар | Ангиотензин I (ангиотензин II ге айналады) |
| Простагландиндер А1 және А2 | |
| Өкпеде метаболизденбейтін заттар | Адреналин |
| Гистамин | |
| Дофамин | |
| Изопротеренол | |
| Простагландиндер |
Өкпенің ең жақсы зерттелгені- биогенді аминдер, соның ішінде бірінші орында серотониннің метаболизмдегі рөлі толықанықталған. Оқшауланған өкпе перфузияланған (перфузанттан) сұйықтықтан, Na+ иондарының градиен– тінен түзілген энергияның нәтижесінде, активті тасымалдау арқылы серотонинді тез сорып алады. Өкпенің серотонинді ұстап алуына қарсы конкурентті ингибиторлар, мысалы, жасушалық тынысалу ингибиторлары және Nа+, K+ - АТФаза – уабаин ингибиторы оны тежейді (8 сурет).
8 сурет. Теңіз шошқасының перфузиялық әдіспен оқшауланған өкпесіндегі сұйықтықтан серотонинді шығаруы. Логарифмдік серотонин концентрациясының перфузия уақытына тәуелділігі берілген. Имипрамин (аминдерді тасымалдаудағы конкурентті ингибитор). KCN (тынысалу ингибиторы), натрий йодоацетаты (гликолиз ингибиторы) және уабаин (Nа
+, K+ - АТФАза ингибиторы) перфузанттан серотониннің шығуын тежейді .
Қандағы өкпемен жұтылып алынған серотонин моноаминооксидаза ферментімен тотығып 5-гидроксииндолсірке қышқылын түзеді. Бұл реакцияның жылдамдығы серотониннің өкпеге жұтылуына қарағанда жылдамырақ болады, сондықтан серотонинді қанағысынан бөлуде лимиттейтін саты осы аминді өкпемен ұстап алу сатысы болып табылады. Авторадиографиялық зерттеу нәтижесінде серотонинді жұтып алуда маңызды рөлді эндотелий атқаратыны көрсетілген. Серотонинге қарағанда басқа биогенді амин – гистамин, қаннан өкпеге жұтылмайды және керекті жылдамдықта метаболизденбейді. Өкпе норадреналинді жұтады және метаболизмдейді,бірақ адреналин жұтылмайды және метаболизмге ұшырамайды. Сонымен, өкпе биогенді аминдерді жұтуда және олардың метоболизмдерінде арнайылықты әсер көрсетеді. Өкпе БАЗды қаннан жұтатын және тотықтыратын , солардың нәтижесінде вазобелсенді қосылыс- тарды өзгертетін тек бір ғана механизм емес.
Басқа механизмнің табиғаты полипептидті БАЗды эндотелий бетінде жинақталған арнайылықты протеазалармен протеолиттік ыдырату болып табылады. Шектеулі протеолиз арқылы активсіз ангиотензин I активті ангиотензин II.ге айналады. Протеолитикалық ыдырату жолымен басқа вазоактивті пептид – брадикинин активсізденеді. Биологиялық активті пептидтерді өкпеде протеолитикалық модификациялау қаннан жұтылуды керек етпейді , процесс эндотелий бетінде жүреді.
Өкпеде жүретін протеолиз процесі туралы айтқанда , қан плазмасындағы альбуминнің фракциясы бар нәруыздардың маңызды рөлін еске алу керек. Олар протеаза α1-антитрипсин ингибиторы, α1-антитрипсин серин протеазаларына жататын тұқымдас ингибитор, олар серпиндер деп аталады. α1-антитрипсин қанға бауырдан секреттеледі. Бұл нәруыздың маңызды физиологиялық рөлі өкпеде іске асады және оның жұмысы ауа тасымал- данатын жол қуысында нейтрофильді лейкоциттермен секреттелетін эластаза ферментінің тым шектен шығатын әсерін тежейді . Эластин белогын қорыту кезінде фермент өкпе тіндеріндерін бұзуы мүмкін. α1-антитрипсиннің туа біткен жетіспеушілігінен, өкпе тіндерінің созылғыштық қасиеттерінің төмен- деуінен, ересектерде эмфизема дамуы байқалады. Эмфизема – альвеола қабырғаларының бұзылуынан өкпе ауруының тууы .
Альфа-1 антитрипсин бауырда синтезделеді. Электрофорез кезінде оның алу орны альфа-1 глобулин фракциясына сәйкес келеді. Қалыпты жағдайда өкпеде фермент –