ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 27.11.2019
Просмотров: 4047
Скачиваний: 98
"включать" липоксигеназный каскад в хлоропластах. Эти
вопросы составляют лишь часть проблемы участия хлоро-
пластов в функционировании липоксигеназнои сигнальной
системы и общей сигнальной сети клеток растений.
В настоящее время имеется достаточно убедительная
информация, чтобы считать липоксигеназный путь превра-
щения мембранных липидов самостоятельной сигнальной
системой. Одним из признаков сигнальных систем является
не только передача сигнала в генетический аппарат клеток,
но и его значительное усиление (принцип фотоумножи-
теля). Взаимодействие одной исходной сигнальной молеку-
лы с рецептором может привести к появлению миллионов
молекул, определяющих ответную реакцию клетки. Так же
как в других сигнальных системах, в липоксигеназнои взаи-
модействие первичного сигнала с рецептором плазмалеммы
активирует фермент (фосфолипазу А
2
), обеспечивающий
передачу информации по сигнальной цепи. Накопление сво-
бодных линолеата или линолената (субстратов липоксигеназ),
вызванное активацией фосфолипазы А, приводит к экс-
прессии генов липоксигеназ [Veronesi et al., 1996], активируя
тем самым липоксигеназную сигнальную систему.
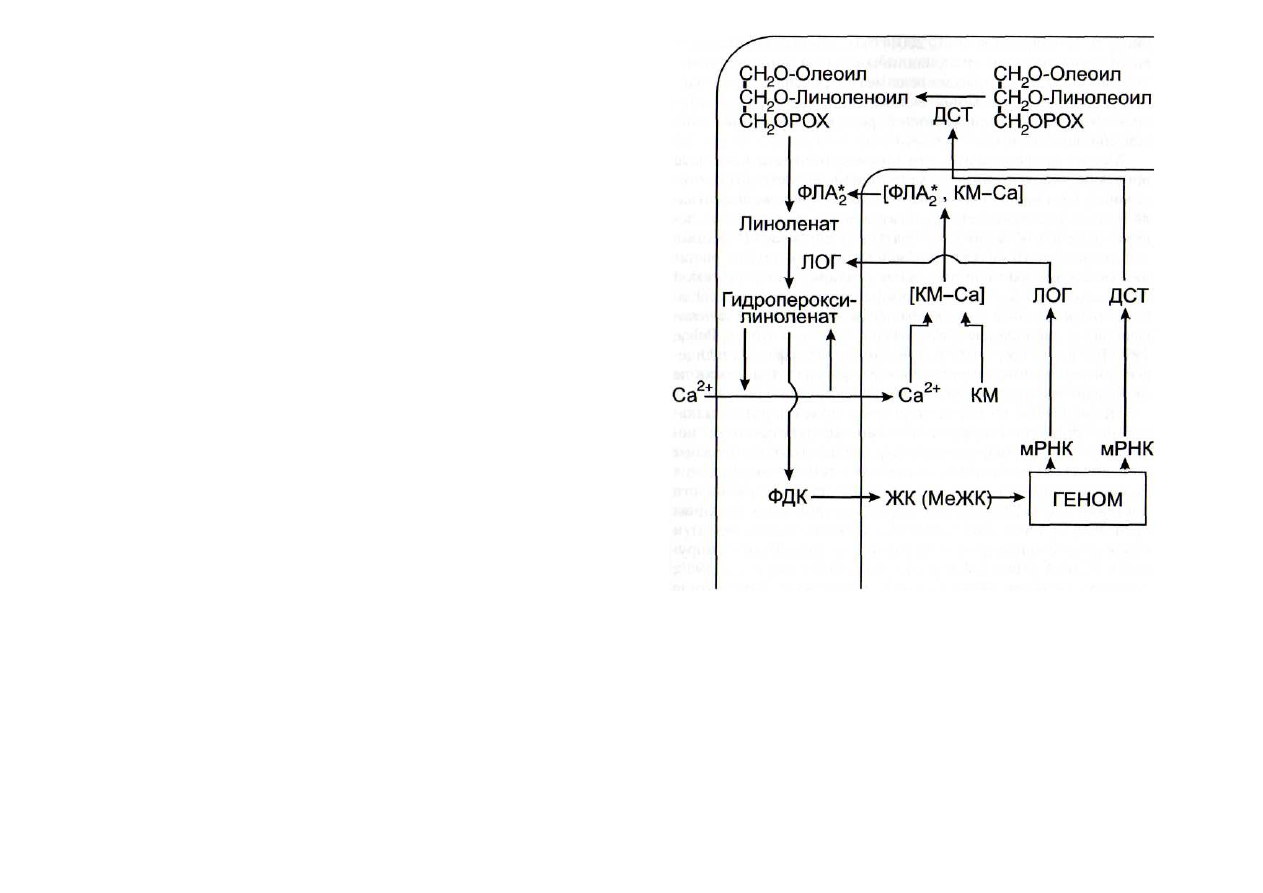
Одной из особенностей усиления сигналов в липоксиге-
назнои системе является использование нескольких видов
автокаталитических процессов (циклов). В частности, это -
автокаталитическое усиление сигнала с участием ионов
кальция и кальмодулина (рис. 21) [Leshem, 1987]. Образую-
щиеся в плазмалемме из линолената или линолеата гидро-
пероксиформы этих кислот могут выступать в роли ионо-
форов, переносящих ионы кальция снаружи внутрь клетки
по градиенту концентрации (известно, что концентрация
ионов кальция за пределами плазмалеммы на 2-3 порядка
выше, чем в цитозоле). Повышение концентрации ионов
кальция в цитозоле приводит к активации фосфолипаз А
при участии кальмодулина и вследствие этого - к еще боль-
шему освобождению полиеновых жирных кислот из фос-
фолипидов. Второй механизм усиления липоксигеназного
метаболизма - это опосредованная жасмонатом [Jensen et
al.,
1997] или метилжасмонатом [Bell, Mullet, 1991; 1993;
Melan et al., 1993; Geerts et al., 1994; Eiben, Slusarenko,
1994; Avdiushko et al., 1995; Veronesi et al.,
1996] индукция
экспрессии генов липоксигеназ (см. рис. 21), приводящая к
Рис. 21. Автокаталитические реакции в липоксигеназнои сиг-
нальной системе [Гречкин, Тарчевский, 1999]
ДСТ - десатураза; ЖК - жасмоновая кислота; КМ - кальмодулин;
| КМ-Са] - комплекс кальмодулин-Са; ЛОГ - липоксигеназа; МеЖК -
метилжасмонат; ФДК - фитодиеновая кислота; ФЛА
2
-
фосфолипаза А
2
повышению скорости оксигенирования линолеата и лино-
лената. Третий автокаталитический цикл - индукция ме-
тилжасмонатом экспрессии генов десатуразы, катализиру-
ющей превращение линолевой кислоты в линоленовую
[ Nishiuchi et al.,
1997]. По всей вероятности, это самый про-
тяженный автокаталитический оксилипиновый цикл (см.

рис. 21). Недавно [Seo et al., 2001] был обнаружен еще один
авто каталитический цикл, заключающийся в индукции ме-
тилжасмонатом экспрессии гена метилтрансферазы (S-аде-
нозил-метионин: жасмоновая кислота - карбоксил-метил-
трансферазы), катализирующей реакцию метилирования
жасмоновой кислоты.
Можно предположить, что промежуточные и конечные
продукты липоксигеназного метаболизма активируют проте-
инкиназы и, таким образом, осуществляют умножение сигна-
ла и его передачу на геном растительных клеток. К сожале-
нию, сведений об этом в литературе очень мало. Известно,
что активировать мембраносвязанные протеинкиназы расте-
ний способны оба типа продуктов реакции, катализируемой
фосфолипазами А
2
, -
как лизофосфолипиды (особенно, лизо-
фосфатидилхолин и лизофосфатидная кислота), так и нена-
сыщенные жирные кислоты [Scherer, 1996 a, b; Klucis, Polya,
1987; Lucantoni, Polya,
1987]. При этом лизофосфатидилглице-
рол, лизофосфатидилсерин и лизофосфатидилэтаноламин не
оказывают активирующего действия на протеинкиназы.
Несмотря на то что конкретные молекулярные меха-
низмы активации генов различными оксилипинами еще не-
достаточно изучены, можно утверждать, что они обладают
способностью вызывать экспрессию генов, кодирующих
белки, принимающие участие в повышении устойчивости
растений к абиогенным стрессорам и в защитных реакциях
против патогенов. Так, экзогенный жасмонат приводит к
синтезу целого набора так называемых жасмонатиндуциру-
емых белков [Mueller-Uri et al., 1988; Herrmann et al., 1989;
Sembdner, Parthier
1993]. Среди них имеются ингибиторы
протеиназ 1 и 2, ингибитор трипсина, тионин, напин, круци-
ферин, вегетативные запасные белки, фенилаланин-аммо-
ний-лиаза, халконсинтаза, липоксигеназа, полифенолокси-
даза и др. Имеются данные [Farmer, Ryan, 1992], что не толь-
ко жасмонат, но и его метаболические предшественники -
13(8)-
гидроперокси-9(г),(Е),15(Х)-октадекатриеновая кис-
лота и 12-оксо-10,15(г)-фитодиеновая кислота - иницииру-
ют образование стрессовых белков, в частности ингибито-
ров протеиназ. Высказывается мнение [Parchmann et al.,
1997], что 12-оксофитодиеновая кислота, проявляющая да-
же большую активность в индукции защитных белков, чем
жасмонат, является главным индуктором экспрессии генов
при местной реакции клеток на стрессоры, а за счет жасмо-
ната и метилжасмоната обеспечивается передача сигнала в
удаленные от места воздействия стрессора ткани и органы
и формирование в них системного иммунитета. Это мнение
подтверждается тем, что в отличие от жасмоната 12-оксо-
фитодиеновая кислота не секретируется из культуры расти-
тельных клеток в среду.
Обнаружено также, что летучие продукты липоксигеназ-
ного метаболизма могут индуцировать (в том числе в сосед-
них растениях) защитную реакцию. Так, т/?анс-2-гексеналь
вызывал образование фенилаланин-аммоний-лиазы, катали-
шрующей образование предшественников растительных ан-
тибиотиков - фенилпропаноидных фитоалексинов. Показа-
но [Fukuda et al., 1997], что 4-гидрокси-2-ноненаль
индуциро-
вал
синтез глутатион-Б-трансферазы, участвующей в обез-
вреживании токсичных для растений веществ.
Одно из свойств сигнальных систем - возможность тор-
можения (прерывания) прохождения сигнала после того,
как он был воспринят клеткой. Такая возможность была
продемонстрирована и в отношении липоксигеназной сиг-
нальной системы. Установлено [Avdiushko et al., 1995], что
метилжасмонат активирует 13-гидропероксидлиазу; это
приводит не только к усилению продукции бактерицидных
и фунгицидных гексеналей, но соответственно к уменьше-
нию доли метаболического потока, направляемого в сторо-
ну жасмоната и метилжасмоната (эффект автоингибирова-
н ия своего синтеза, проявляющийся, по-видимому, при дос-
тижении пороговых концентраций метилжасмоната). Ока-
залось, что дивиниловые эфиры - колнелевая и этеролевая
кислоты - могут ингибировать активность липоксигеназ и
таким образом затруднять функционирование липоксиге-
пазной сигнальной системы [Corey et al., 1987]. Колнелевая
кислота является активным ингибитором 9-липоксигеназы,
j-
геролевая кислота - 13-липоксигеназы [Гречкин, неопуб-
ликованные данные]. Предполагается, что ингибирующее
действие на липоксигеназную активность может также ока-
зывать эпоксидный продукт превращения 9-гидроперокси-
линолената [Sok, Kim, 1989].
Итак, результаты изучения сигнальных функций липок-
сигеназного метаболизма позволяют считать, что ему при-
сущи основные свойства, характерные и для других сиг-
нальных систем, - рецепция, преобразование и умножение
сигнала, приводящие (при участии протеинкиназ) к экспрес-
сии определенных генов и соответствующему ответу расти-
тельной клетки.
В связи с тем что участие протеинкиназ является важ-
нейшим звеном сигнальных систем клеток растений, осо-
бую значимость приобретают данные о влиянии интермеди-
атов липоксигеназного метаболизма на фосфорилирование
белков растений [Каримова и др., 19996]. Показано, что эк-
зогенная 12-гидроксидодеценовая кислота (ГДК) в неболь-
ших концентрациях является очень активным стимулято-
ром роста, более активным, чем травматин и жасмоновая
кислота [Гречкин, 1992]. Поскольку нельзя было исклю-
чить возможность участия ГДК в функционировании сиг-
нальных систем, мы поставили перед собой задачу исследо-
вать ее влияние на фосфорилирование белков, имея в виду,
что протеинкиназные реакции являются важнейшим зве-
ном всех известных сигнальных систем клеток.
ГДК в широком диапазоне концентраций (с максимумом
при 10"
7
М) вызывала значительное повышение содержа-
ния радиоактивного фосфата во фракции растворимых бел-
ков. Поскольку при гомогенизации листьев и в реакцион-
ной смеси использовались неспецифические ингибиторы
фосфатаз, эффект отщепления фосфата от белков в про-
цессе обработки материала был минимальным. ГДК оказы-
вала больший стимулирующий эффект на фосфорилирова-
ние белков, чем такой известный эффектор протеинкиназ-
ной активности, как цАМФ. Метилжасмонат также вызы-
вал большее фосфорилирование белков, чем цАМФ, но
меньшее, чем ГДК.
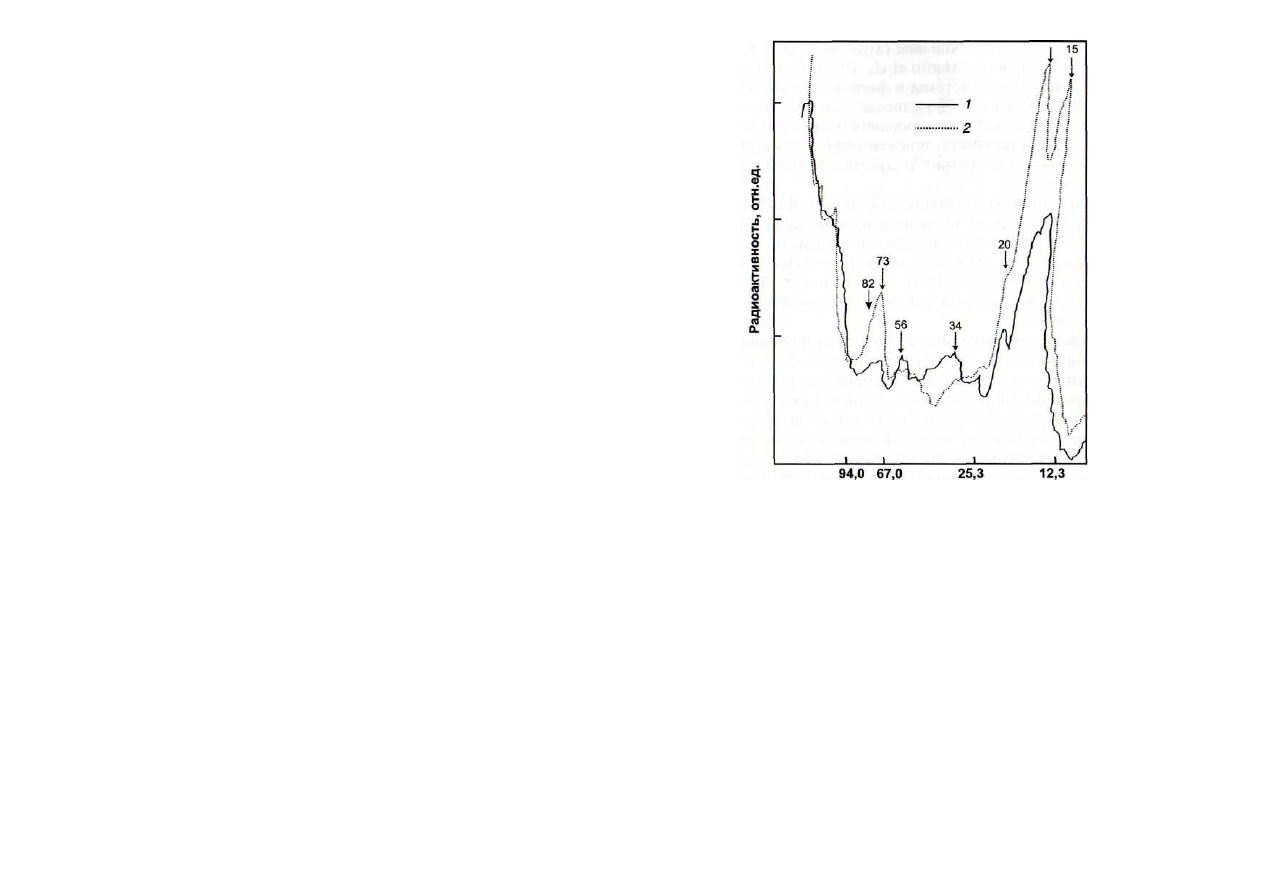
Результаты экспериментов по разделению фосфорили-
рованных белков и определению их радиоактивности пока-
зали, что обработка ГДК приводила к значительному повы-
шению уровня фосфорилированности трех полипептидов
низкой молекулярной массы (10-24 кДа) и высокомолеку-
лярного полипептида 70 кДа (рис. 22). Из данных рисунка
следует, что уровень радиоактивности отдельных полипеп-
тидов (в области 40 кДа) был ниже в варианте с обработкой
растений ГДК.
Фосфорилирование белков участвует в регуляции мно-
жества процессов [Protein Phosphorylation in Plants, 1996].
Молекулярная масса белков, кДа
Рис. 22. Влияние 12-гидроксидодеценовой кислоты (ГДК) на фос-
форилирование белков [Каримова и др., 19996] гомогената
трехдневных растений гороха
1
-
контроль; 2 - ГДК (0,1 мкМ). Рентгенограммы сканировали на
денситографе ИФО-450. Стрелками указаны молекулярные массы бел-
ков, радиоактивность которых подвергалась наибольшим изменениям
Получены данные о существовании в растениях различных
типов протеинкиназ и об участии фосфорилирования бел-
ков в экспрессии защитных генов после взаимодействия
различных патогенов, элиситоров и стрессовых гормонов с
клетками растений [Grab et al., 1985; Dietrich et al., 1990;
Farmer et al., 1991; Bolwell et al.,
1996]. О важной роли фос-
форилирования белков в формировании устойчивости рас-
тений к патогенам свидетельствуют данные о том, что не-
которые клонированные гены, кодирующие защитные бел-
ки растений, повышающие их устойчивость к инфицирова-

нию патогенами, имели большое сходство с генами, кодиру-
ющими протеинкиназу [Martin et al., 1993]. Если роль в це-
лом липоксигеназной системы в формировании защитного
ответа растений достаточно известна, то данные о значи-
тельном усилении фосфорилирования белков под влиянием
промежуточного продукта липоксигеназного метаболизма
(самой быстрой его ветви) получены [Каримова и др.,
19996] впервые.
Тот факт, что под влиянием ГДК уровень фосфорилиро-
ванности различных полипептидов изменился в разной сте-
пени (см. рис. 22), свидетельствует в пользу того, что она
может неодинаково влиять на активность ферментов (про-
теинкиназ и протеинфосфатаз), определяющих баланс ско-
ростей фосфорилирования и дефосфорилирования отдель-
ных полипептидов.
Повышение уровня общего фосфорилирования белков
под влиянием цАМФ не вызывает удивления, так как из-
вестно существование цАМФ-активируемых протеинки-
наз. Большее фосфорилирование белков под влиянием ме-
тилжасмоната тоже объяснимо, так как известно, что жас-
монат и метилжасмонат могут включать (пусть не полно-
стью) ответную реакцию растений, характерную для меха-
нического повреждения тканей или инфицирования пато-
генами. Известно, что в этих случаях активируются сиг-
нальные системы клеток, в том числе не только циклоаде-
нилатная. Наибольшее фосфорилирование белков под вли-
янием ГДК свидетельствует или о существовании активи-
руемых непосредственно ГДК протеинкиназ, или о вклю-
чении с помощью экзогенной ГДК совокупности сигналь-
ных систем клеток, причем не только циклоаденилатной
(поскольку цАМФ вызывал меньший эффект), но и каль-
циевой, и НАДФ-оксидазной [Chandra, Low, 1995], и, воз-
можно, "собственной" липоксигеназной системы. Факты
такого влияния продуктов фосфолипазных и липоксиге-
назных реакций обнаружены у животных [Гамалей, Клю-
бин, 1996] и растительных [Гречкин, Тарчевский, 1999]
клеток.
Выявлено [Гречкин и др., 1987], что ГДК является пре-
обладающим продуктом метаболизации гидроперекисей
линолевой и линоленовой кислот у бобовых. Высокая рост-
стимулирующая активность 12-ГДК обнаружена в опытах
Рис. 23. Схема радиоавто-
графов электрофореграмм
белков, характеризующая
влияние
инфицирования
микоплазмами и обработки
жасмоновой кислотой на
синтез полипептидов [Тар-
чевский и др., 19966]
1
-
контроль; 2 - инфици-
рование микоплазмами;
3
-
об-
работка жасмоновой кислотой.
Слева от столбика 1 - мо-
лекулярные массы маркерных
белков
с каллусной культурой
сои [Гречкин, 1992; Гречкин и др., 1987].
Как известно, уровень фосфорилированности белков
определяется активностью не только приходной протеин-
киназной, но и расходной протеинфосфатазной реакции. В
растениях обнаружены высокоактивные протеинфосфата-
зы нескольких типов [MacKintosh, Cohen, 1989; Vera-Estrella
et al., 1994; Xing et al.,
1996], которые могут контролиро-
ваться с помощью различных эффекторов. Насколько
большой вклад вносит активность фосфатаз в изменение
уровня фосфорилированности полипептидов под влиянием
ГДК, остается невыясненным.
Изменение спектров фосфорилированности белков яв-
ляется достаточно важным показателем активности интер-
медиатов сигнального пути. Чаще изучается их влияние на
синтез белков. Мы исследовали образование жасмонатин-
дуцированных белков в сопоставлении с влиянием на этот
процесс инфицирования растений микоплазмами Achole-
plasma laidlawii
118 [Тарчевский и др., 19966].
Результатом действия на растения экзогенной жасмоно-
вой кислоты (5 • К)-
5
М) было увеличение поглощения
14
С-лей-
цина и включения радиоактивной метки в белки. Относи-
тельная доля включения
14
С в белки от поглощения также
возрастала. При разделении полипептидов с помощью элек-
трофореза было обнаружено индуцированное жасмонатом
образование двух новых полипептидов - 38 и 42 кДа (рис. 23).
Если учесть, что появление нового, индуцированного
жасмонатом белка 38 кДа наблюдается и при инфицирова-
нии микоплазмами, то можно сделать вывод, что заражение
растений включает классический, характерный для биоген-
ного стресса, катаболический липидный сигнальный путь:
активация фосфолипазы А
2
—
> освобождение линолената из
фосфолипидов мембран —» липоксигеназное превращение
его в 13-пероксилиноленат -> образование жасмоновой ки-
слоты в результате гидропероксидциклазной и сопутствую-
щих реакций [Vick, Zimmerman, 1987; Гречкин, 1992; Тар-
чевский, 1993] —
>
активация генов устойчивости —> образо-
вание жасмонатиндуцированных белков —» формирование
местной и системной устойчивости к патогенам [Neumann et
al., 1989; Sembdner, Parthier, 1993].
Так как жасмонат вызывал индукцию образования лишь
одного из трех микоплазмаиндуцированных белков, то
можно предположить, что он является только частью сиг-
нальной системы, приводящей к формированию иммуните-
та растений [Тарчевский и др., 19966].
НАДФН-ОКСИДАЗНАЯ СИГНАЛЬНАЯ СИСТЕМА
Вызванная действием элиситоров на клетки растений
активация НАДФН-оксидазы - стартового фермента
НАДФН-оксидазной системы (рис. 24), является причиной
интенсивного образования активных форм кислорода - сво-
бодных радикалов О
2
и OFT, а также перекиси водорода.
Впервые такой окислительный "взрыв" наблюдали в 1983 г.
[Doke et al.,
1996] в ответ на инфицирование клубней карто-
феля фитофторой. Позднее обнаружили появление окисли-
тельного "взрыва" под влиянием различных патогенных
грибов, бактерий, вирусов, элиситоров, механического по-
вреждения растений.
Обращает на себя внимание преходящий характер окис-
лительного "взрыва", проявляющийся в относительно бы-
стром возврате концентрации перекиси водорода к исход-
ному уровню (рис. 25). Это может быть вызвано как сниже-
нием активности НАДФН-оксидазы, в результате чего
уменьшается приходная часть баланса перекиси водорода,
так и усилением расходной части этого баланса, а именно
выходом части Н
2
О
2
за пределы клеток (где она использу-
ется в пероксидазных реакциях), разрушением перекиси во-
дорода каталазой [Willekens et al., 1997], потреблением
части Н
2
О
2
в аскорбат-пероксидазной реакции аскорбат-
глутатионового цикла (рис. 26).
В образование активных форм кислорода при действии
элиситоров, кроме НАДФН-оксидазы, могут вносить вклад
и другие ферменты, например локализованные в клеточной
стенке оксалатоксидаза [Zhou et al., 1998] и пероксидазы,
активирующиеся при щелочном сдвиге рН за пределами
плазмалеммы, который наблюдается при инфицировании
патогенами и действии элиситоров [Wojtaszek, 1997]. При
этом была обнаружена патогениндуцированная секреция