ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 27.11.2019
Просмотров: 4041
Скачиваний: 98
ков, а от них - на фосфолипазу С (ФЛС), катализирующую
реакцию гидролиза эфирной связи между остатками фос-
форной кислоты и гидроксила глицерина фосфоинозитоль-
ного фосфолипида - фосфатидилинозитолбисфосфата
(ФИФ). Образующиеся диацилглицерин и инозитол-1,4,5-
трисфосфат являются вторичными посредниками. Первый
может активировать мембранные Са
2+
-
зависимые проте-
инкиназы С (ПКС). Изоформы этого фермента отличают-
ся различной степенью активации ионами кальция и диа-
цилглицерином. Протеинкиназы С способны осуществлять
фосфорилирование большого числа белков, регулируя их
активность и вызывая клеточный ответ на внешний сигнал.
Это относится и к фосфорилированию белковых факторов
регуляции транскрипции.
В растениях существует несколько изоформ ФЛС. Наи-
больший интерес представляют две из них. Первый тип
ФЛС связан с поверхностью плазматических мембран, ее
субстратами являются полифосфоинозитиды, необходимая
концентрация свободных Са
2+
находится в физиологиче-
ской области (от 1 нМ до 1 мкМ); второй тип ФЛС - преи-
мущественно растворимая ФЛС, в качестве субстрата для
нее предпочтителен фосфатид ил инозитол, для полной ак-
тивации фермента необходима высокая концентрация сво-
бодных ионов Са
2+
(мМ) [Drobak et al., 1996]. Имеются не-
прямые доказательства об участии G-белков в индукции
гидролиза фосфатидилинозитолбисфосфата.
Другой вторичный посредник - инозитол- 1,4,5-трисфос-
фат, взаимодействует с белками кальциевых каналов тоно-
пласта и эндоплазматической сети и открывает их, что вы-
зывает поток ионов кальция в цитозоль (рис. 13). В нем Са
2+
активирует различные ферменты, например кальцийзави-
симые протеинкиназы (ПКС) или кальций-кальмодулинзави-
симые протеинкиназы (ПКВ), которые, в свою очередь, мо-
гут фосфорилировать белки, в том числе факторы регуля-
ции транскрипции, и вызвать экспрессию защитных генов.
Инозитол-1,4,5-трисфосфат (или продукт его фосфорили-
рования - инозитолтетракисфосфат) может повышать кон-
центрацию Са
2+
в цитозоле, открывая также кальциевые
каналы плазмалеммы.
Предполагается, что у животных клеток мономерные
трансмембранные белки - рецепторы упомянутых выше
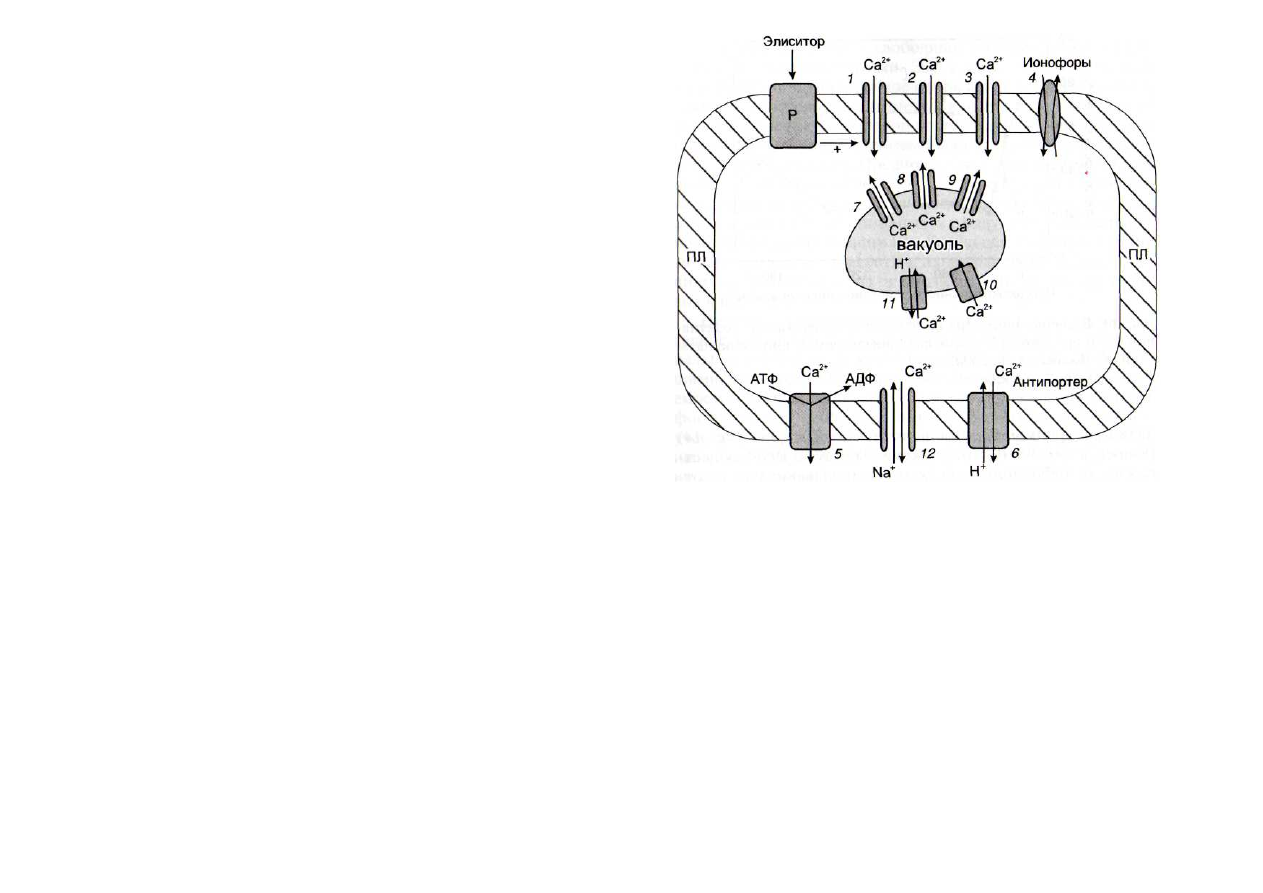
Рис. 13. Влияние элиситоров на кальциевый обмен клеток расте-
ний
1 -
рецепторактивируемый Са
2+
-
канал; 2 - кальциевые каналы, ак-
тивируемые ИФ
3
и ИФ
4
;
3 -
потенциалзависимые кальциевые каналы;
4 -
транспортировка ионов Са
2+
ионофорами; 5 - Са
+
-
АТФазы плазма-
леммы;
6 -
Са /Н
+
-
антипортер плазмалеммы; 7 - кальциевые каналы,
активируемые ИФ
3
;
8
-
кальциевые каналы, активируемые цАДФР; 9 -
кальциевые каналы, активируемые протеинкиназами;
10
-
Са
2+
-
АТФазы
тонопласта;
11 -
Са
2+
/Н
+
-
антипортер тонопласта;
12 -
Ca
2+
/Na
+
-o6-
менник; ПЛ - плазмалемма; Р - рецептор
инозитолфосфатов, после взаимодействия с ними образуют
тетрамерные каналы, осуществляющие вброс ионов каль-
ция в цитозоль [Крутецкая, Лебедев, 2001].
Недавно обнаружено новое семейство кальцийзависи-
мых протеинкиназ (КЗПК), отличных от ПКС. Показано,
что под влиянием элисторов может происходить вызван-
ная фосфорилированием фермента трансформация его не-

Продолжительность действия элиситора, мин
Рис. 14. Влияние элиситора на изменение содержания неактив-
ной (7) и активной (2) форм кальцийзависимой протеинкиназы
(КЗПК) [Romeis et al., 2000] 7-70 кДа; 2-68 кДа
активной формы 68 кДа в активную 70 кДа (рис. 14)
[Romeis et al.,
2000]. Предложена [Trewavas, 1999] сущест-
венная модификация этой схемы, основывающаяся на от-
носительно медленном передвижении ионов кальция в ци-
тозоле, как было показано в опытах с использованием
инъекции меченого кальция в гигантский аксон кальмара.
Причинами могло быть интенсивное связывание ионов
кальция белками и обратный перенос избытка ионов каль-
ция Са
2+
-
активируемыми АТФазами. Новая схема распро-
странения кальциевой волны в клетках предполагает, что
после открывания кальциевого канала у его отверстия
происходит накопление относительно медленно диффун-
дирующих ионов кальция, что вызывает активацию в этой
области мембраносвязанной фосфолипазы С. Освобожда-
ющийся в результате фосфолипазной реакции инозитол-
трисфосфат подвижен и, диффундируя от места образова-
ния, может достигать соседних кальциевых каналов, свя-
зываться с ними и открывать их. Необходимо иметь в ви-
ду, что белки каналов имеют места связывания не только
ИФ
3
, но и ионов кальция. Предполагается, что при локаль-
ном передвижении от соседнего открытого кальциевого
канала они достигают свободных кальцийсвязывающих
мест и захватываются ими. Это вносит дополнительный
вклад в ИФ
3
-
индуцированное открывание и поддержание в
открытом состоянии кальциевых каналов. Так происходит
распространение кальциевой волны вдоль мембраны и од-
новременно, местное (примембранное) повышение содер-
жания ионов кальция. Предполагается, что этот механизм
проявляется в том случае, когда концентрация элиситора
невелика и лимитирует количество активируемых кальци-
евых каналов. Необходимо иметь в виду, что значительное
повышение концентрации ионов кальция в цитозоле вбли-
зи каналов может привести к их закрыванию и ограниче-
нию поступления Са
2+
из окружающей среды или органои-
дов в цитозоль.
Передача элиситорного сигнала в геном клеток, интен-
сивность и направленность функционирования этой сиг-
нальной системы осложнена различными деталями, касаю-
щимися природы элиситоров, большей или меньшей атаку-
е мости фосфолипазой С различных молекулярных видов
фосфолипидов, особенностями строения изоформ белков -
участников сигнальной системы, различиями вклада каль-
циевых каналов плазмалеммы и различных органелл клет-
ки в кальциевую "вспышку", наконец, вероятной "класте-
ризацией" кальциевых каналов и кальцийзависимых Са
2+
-
АТФаз и удаленностью друг от друга этих кластеров
|Trewavas,
1999]. Недавно была высказана гипотеза [Олов-
ников, 2001] о существовании во внутренней ядерной мемб-
ране животных клеток кластеров специальных, например
кальциевых каналов, с помощью которых осуществляется
локальное (фонтанное) изменение концентрации ионов
вблизи определенных генов и таким образом происходит
специфическая регуляция их экспрессии. Топографическая
специфичность регуляции генов могла бы осуществляться с
помощью специальной фонтанной РНК (фРНК) и так на-
зываемых фионов - участков ДНК, способных связывать
фРНК. Вброс порции ионов в ядро происходит с помощью
комплекса фион-фРНК-белок ионного канала внутренней
мембраны ядерной оболочки.
Имеется ряд обзорных работ [Gilroy et al., 1993; Poovaiah,
Reddy, 1993; Bush, 1995; Trewavas, Malho,
1997; Ткачук, 1998;
Sanders et al., 1999; Trewavas, 1999; Bowler, Fluhr, 2000; White,

2000; Reddy,
2001], посвященных сигнальной функции
ионов кальция, в которых анализируются особенности
функционирования структур, обеспечивающих как повы-
шение концентрации ионов кальция в цитозоле (кальцие-
вые каналы), так и снижение - до исходного уровня с помо-
щью связывания избытка ионов кальция белками, разруше-
ния (дефосфорилирования) ИФ
3
и вследствие этого закры-
вания кальциевых каналов, а также с помощью ионных
помп, перебрасывающих ионы кальция обратно против гра-
диента концентрации за счет использования энергии гидро-
лиза макроэргических фосфатных связей АТФ (см. рис. 13).
У высших растений охарактеризованы различные
Са
2+
-
каналы, по которым Са
2+
транспортируется через
плазматические мембраны, тонопласт, мембраны эндоплаз-
матической сети, хлоропластов и ядер [White, 2000]. Эти ка-
налы подразделяются на несколько групп, в зависимости от
их электрических характеристик. Они в разной степени чув-
ствительны к верапамилу и La
3+
.
В функционирование сиг-
нальных путей вовлечены главным образом, кальциевые
каналы, активируемые деполяризацией мембран от -140 мВ
до менее отрицательных значений, что, по-видимому, при-
водит к изменению конформации белков кальциевых кана-
лов и их открыванию. Элиситор-активируемые каналы вы-
делены в отдельную группу [White, 2000].
ИТФ
3
-
и цАДФрибоза-управляемые каналы найдены в
мембранах ЭПР и вакуолей растений, тогда как в клетках
животных ИТФ
3
и цАДФрибоза индуцируют выход Са
2+
только из ЭПР. Обнаружено, что разные типы стрессоров
индуцируют выход Са
2+
в цитозоль из разных внутрикле-
точных компартментов [Reddy, 2001].
Последующее за "кальциевой вспышкой" снижение кон-
центрации Са
2+
в цитозоле является обязательным условием
функционирования кальциевой сигнальной системы. Более
того, длительное сигналиндуцированное повышение концен-
трации ионов кальция может привести к гибели клеток.
Существует несколько механизмов понижения уровня
Са
2+
в цитозоле. Оно может осуществляться за счет связы-
вания Са
2+
кальмодулином и другими белками. Са
2+
-
связывающие белки, обнаруженные в растениях, под-
разделяются на четыре группы: 1) кальмодулин (КМ);
2) КМ-подобные белки с кальцийсвязывающими доменами;
3) Са
2+
-
регулируемые протеинкиназы; 4) белки без специ-
фического Са
2+
-
связывающего домена.
У животных клеток одна молекула кальсеквестрина, не
имеющая такого домена, связывает до 43 ионов кальция за
счет их взаимодействия с остатками аспарагиновой и глута-
миновой кислот. Каждая молекула другого активного бел-
ка - кальретикулина, связывает ионы кальция с помощью
специального домена [Крутецкая, Лебедев, 2001].
Привлекают все большее внимание аннексины -Са
2+
-
связывающие белки, взаимодействующие с кислыми
фосфолипидами в присутствии Са
2+
. Некоторые аннексины
способны образовывать ионные каналы в искусственных
мембранах [Минкин и др., 1998]. Показано, что аннексины
Arabidopsis thaliana
участвуют в защите от окислительного
стресса [Gidrol et al., 1996].
Пожалуй, основной вклад в снижение концентрации
ионов кальция в цитоплазме играют закрывание кальциевых
каналов в результате гидролиза ИФ
3
специфическими фос-
фатазами и активация кальциевых насосов (Са
2+
-
АТФаз),
которые за счет энергии гидролиза АТФ переносят ионы
кальция в обратном направлении против градиента концен-
трации, восстанавливая исходные значения градиента Са
2+
и
is
связи с этим способность клеток воспринимать новый эли-
ситорный сигнал. Са
2+
-
АТФазы характеризуются высоким
сродством к Са
2+
.
В растениях найдены различные Са
2+
-
АТФазы, принад-
лежащие в том числе к автоингибирующемуся (АСА) типу
(которые регулируются комплексом Са
2+
-
кальмодулин).
АСА-тип Са
2+
-
АТФаз растений локализуются в ЭПР и
плазматических мебранах, тогда как в животных клетках
этот тип АТФаз локализован исключительно на плазмати-
ческих мембранах. Активность АСА-типа Са
2+
-
АТФаз ЭПР
в Arabidopsis ингибируется Са
2+
-
зависимой протеинкиназой |
Hwang et al.,
2000]. Обнаружены зависимые и независимые
от кальмодулина Са
2+
-
АТФазы. Установлен элиситоринду-
цированный синтез кальмодулин-стимулируемой кальцие-
вой АТФазы плазмалеммы [Chung et al., 2000].
Еще один механизм снижения содержания ионов каль-
ция в цитозоле - их удаление в процессе работы Са
2+
/Н
+
ан-
типортеров, использующих для этого энергию гидролиза
АТФ. Роль кальциевых каналов и кальциевых помп в мем-
бранах клеток растений была экспериментально обоснова-
на опытами с использованием специфических ингибиторов
[Scheel, 1998].
Возникает вопрос, существует ли в растениях еще один
механизм удаления Са
2+
из цитозоля, характерный для кле-
ток животных после их возбуждения, с помощью
Na
+
/
Са
2+
-
обменника, обладающего низким сродством к
Са
2+
, но высокой скоростью переноса - около 20 нМ на 1 мг
мембранного белка в секунду при 300 °С? Функционирова-
ние этого белка-переносчика осуществляется за счет энер-
гии трансмембранного градиента Na
+
и мембранного потен-
циала. На клетках водорослей получены данные о противо-
положно направленных трансмембранных потоках Са
2+
и
Na
+
,
характеристики которых свидетельствуют о сходстве
их с функциональными характеристиками Na
+
/Ca
2+
-o6MCH-
ника животных клеток [Karimova et al., 2000].
В клетках растений существуют еще два органоида, в
которых концентрация ионов кальция может достаточно
сильно изменяться, - хлоропласты и митохондрии; однако
это в значительной степени автономные образования со
своими системами поддержания ионного гомеостаза. До сих
пор неясно, в какой степени они участвуют в элиситоринду-
цированном изменении концентрации ионов кальция в ци-
тозоле. Есть надежда, что этот вопрос будет разрешен с ис-
пользованием специально сконструированных для этой це-
ли трансгенных растений. Необходимо отметить, что воп-
рос о вкладе митохондрий в функционирование кальциевой
сигнальной системы у животных клеток решается положи-
тельно. Более того, считается, что они принимают актив-
ное участие в сигнальных внутриклеточных процессах
[Крутецкая, Лебедев, 2001], что они могут освобождать
Са в цитозоль с помощью Na
+
/
Са
2+
-
обменника внутренней
мембраны и поглощать, используя Са
2+
-
унинортер.
Для измерения концентрации ионов кальция в цитозоле
и других компартментах используют селективные электро-
ды, красители, а также трансгенные растения с привнесен-
ным геном экворина - Са
2+
-
зависимого флуоресцентного
белка. Использование таких трансгенных растений позво-
лило установить, что сигналиндуцированное преходящее
повышение содержания ионов кальция в цитозоле приводит
к быстрому и преходящему возрастанию их концентрации в
митохондриях, что может быть предотвращено предобра-
боткой разобщителями электронного транспорта и фосфо-
рилирования [Rizzuto et al., 1992]. Установление этого фак-
та позволяет подойти к объяснению до сих пор еще не
очень ясного механизма передачи элиситорного сигнала,
рецептируемого плазмалеммой, в хлоропласты и митохонд-
рии. Получение трансгенных растений с химерным геном
экворина и ядерного белка нуклеоплазмина позволило ус-
тановить, что сигналиндуцированное повышение содержа-
ния Са
2+
происходит не только в цитозоле, но и в ядре [Van
der Luit et al., 1999].
Как уже отмечалось, в целом ряде опытов было показа-
но, что сигналиндуцированное возрастание концентрации
ионов кальция в цитозоле объясняется активацией кальци-
евых каналов не только плазмалеммы, но и внутренних
вместилищ ионов кальция [Knight et al., 1996; Mori et al.,
1998].
В опытах с активатором G-белков мастопараном было
обнаружено, что устранение внешнего пула Са
2+
могло не
ингибировать кальциевого "всплеска" в цитозоле, из чего
был сделан вывод об активации кальциевых каналов мемб-
ран органелл [Takahashi et al., 1998]. Подавление эффекта с
помощью ингибитора неомицина позволило сделать вывод
об участии в этом процессе фосфоинозитидов.
В настоящее время интенсивно обсуждаются возможно-
сти автокаталитических и автосупрессорных процессов в
кальциевой сигнальной системе. Большой интерес вызвали
сообщения о том, что если незначительное повышение кон-
центрации ионов кальция в цитозоле стимулирует, то силь-
ное - ингибирует индуцируемое инозитол-1,4,5-трисфосфа-
том открывание кальциевых каналов. Обнаружен Са
2+
-
ин-
дуцируемый синтез белка кальмодулина, который образует
с ионами кальция комплекс, принимающий участие в акти-
вации различных белков, в том числе протеинкиназ [Romeis
et al.,
1999], и через них - факторов регуляции транскрип-
ции. В растениях существуют и кальмодулин-независимые,
но Са
2+
-
зависимые протеинкиназы, имеющие у С-конца до-
мен, по своей структуре близкий к структуре кальмодулина
и способный связывать ионы кальция, что приводит к акти-
вации протеинкиназы без участия молекулы кальмодулина
| Harper etal., 1991].
В последнее время у животных объектов обнаружена не
только инозитол-1,4,5-трисфосфатная, но еще одна ветвь
кальциевой сигнальной системы - инозитол-3,4,5-трисфос-
фатная, причем предполагается возможность ее функцио-
нирования и в клетках растений [Munnik et al., 1995; 1998].
Оказалось, что изменение местоположения одной из фос-
фатных групп существенно изменяет набор белков, кото-
рые являются мишенями для инозитолтрисфосфата и кото-
рые активируются им.
Возможность регуляции функционирования цитоскеле-
та с помощью кальциевой сигнальной системы - одна из ак-
туальных задач физиологии растений. Имеются данные, по-
зволяющие считать, что действие на микротрубочки и мик-
рофиламенты изменения концентрации ионов кальция опо-
средовано кальмодулином и кальцийзависимыми протеин-
киназами. Динамическое состояние микротрубочек в значи-
тельной степени зависит от фосфорилирования-дефосфо-
рилирования связанных с ними белков. Ингибирование про-
теинкиназ индуцировало стабилизацию микротрубочек в
суспензионной культуре клеток табака при гипотермии. Из-
менения в цитоскелете могут иметь существенные послед-
ствия для клетки, так как от него зависит движение цито-
плазмы, ориентация отложения микрофибрилл целлюлозы
в клеточную стенку [Тарчевский, Марченко, 1987;
Tarchevsky, Marchenko,
1991], транспортные процессы и т.д
Изменения ионных потоков наблюдаются в клетках не
только при действии на них патогенов (и патогенпродуци-
руемых элиситоров), но и при симбиотическом взаимодей-
ствии азотфиксирующих бактерий и бобовых растений, в
результате которого появляются узелки на корнях. Один
из наиболее быстрых ответов растений на действие вызы-
вающих образование корневых узелков (nodules) липохи-
тоолигосахаридов (Nod-факторов) - это повышение в ци-
топлазме концентрации ионов кальция, а также протонов
[Cardenas et al.,
1999; 2000]. Показательно, что кальциевый
ионофор А-23187 по своему действию на образование кор-
невых узелков приближался к эффекту Nod-факторов
[Felle et al., 1998].
Рецепторы Nod-факторов могут быть непосредственно
связаны с Са
2+
и С1-каналами [Cardenas et al., 2000]. Альтер-
нативное мнение заключается в том, что вход кальция - это
следующее звено за гетеротримерным G-белком (о чем сви-
детельствуют опыты с активаторами этих белков мастопа-
раном и пертуссиновым токсином) и фосфолипазой С (опы-
ты с ингибитором фермента - неомицином). В первом слу-
чае наблюдалась индукция, а во втором - ингибирование
эффекта Nod-факторов [Pingret et al., 1998].