ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 27.11.2019
Просмотров: 4039
Скачиваний: 98

СИГНАЛЬНАЯ ФУНКЦИЯ ЦИТОСКЕЛЕТА
От цитоскелета (тубулиновых микротрубочек и актино-
вых микрофиламентов) зависит морфогенез на клеточном
уровне (ультраструктура клеток), осуществление простран-
ственно-временного контроля роста, деления и дифферен-
циации клеток [Васильев, 1996; Клячко, 1998; Медведев,
Маркова, 1998]. Известно, что переориентация расположе-
ния микротрубочек под влиянием фитогормонов приводит
к соответствующему изменению направленности отложе-
ния вновь синтезируемых микрофибрилл целлюлозы [Тар-
чевский, Марченко, 1987; Tarchevsky, Marchenko, 1991] и па-
раметров роста клеток, например, к замедлению или пре-
кращению удлинения клеток и их утолщению. Это может
происходить достаточно быстро. Например, переориента-
ция микротрубочек в замыкающих клетках устьиц осущест-
вляется уже через несколько минут после начала действия
экзогенной абсцизовой кислоты [Eun, Lee, 1997]. Ауксин и
гиббереллины способствуют поперечному расположению
микротрубочек по отношению к продольной оси клеток,
цитокинины, абсцизовая кислота и этилен ориентируют мик-
ротрубочки в продольном направлении. Особенно быстро
клетки реагируют на этилен.
В последнее время обращают все большее внимание на
важную роль сигнальных систем не только в изменении ме-
таболизма, но и в структурных изменениях клеток и участии
в них цитоскелета при патогенезе [Chrispeels et al., 1999;
Volkmann, Baluska, 1999; Papakonstanti et al., 2000].
И у животных, и у растений цитоскелет - сложная сеть
нитей, пронизывающих цитозоль, - состоит из трех основ-
ных типов структур: толстых тубулиновых микротрубочек,
тонких актиновых нитей (филаментов) и промежуточных
(по диаметру) нитей. От этих надмолекулярных образова-
ний, постоянно распадающихся и создаваемых заново из мо-
номерных белков, зависит движение цитозоля, органоидов
и очертания клеток растений, а также внутриклеточный
транспорт ряда белков. Все типы микротрубочек и актино-
вых филаментов способны связываться латерально с помо-
щью вспомогательных белков или с себе подобными или с
другими типами структур. Белки цитоскелета и обслужива-
ющие их белки имеют консервативные последовательно-
сти, характерные и для животных, и для растений (еще один
пример универсальности молекулярной и надмолекулярной
структуры). Микротрубочки образуются в два этапа: снача-
ла путем димеризации а- и Р-тубулина, а затем димеры по-
лимеризуются с образованием трубчатой структуры диа-
метром 22 нм. Вспомогательные МАР-белки (microtubules
associated proteins)
нескольких классов значительно снижа-
ют пороговую концентрацию тубулина, необходимую для
сборки из него микротрубочек. Спиралевидные актиновые
нити диаметром 6-7 нм образуются из однотипных актино-
вых субъединиц с затратой АТФ. Баланс между мономер-
ными актиновыми белками и микрофибриллами обеспечи-
вается вспомогательными белками - актин-деполимеризу-
ющим фактором, кофилином и профилином. С помощью
вспомогательного белка фодрина обеспечиваются лате-
ральное взаимодействие нитей и появление пучков микро-
фибрилл, а вспомогательный белок фил амин участвует в
образовании сетчатой (решетчатой) структуры, скрепляя
филаменты в местах их пересечения друг с другом. Микро-
трубочки и микрофиламенты способны участвовать в пере-
мещении вдоль своей поверхности различных структур
(включая везикулы и органеллы) с помощью специальных
вспомогательных белков-моторов кинезина и цитоплазма-
тического динеина.
Все эти факты приобретают для нас особое значение в
связи с тем, что многие участники процессов функциониро-
вания, образования и деградации микротрубочек и микро-
фибрилл, а также их реориентации являются мишенями ин-
термедиатов некоторых сигнальных систем клеток. Это от-
носится к повышению в цитозоле концентраций протонов,
ионов кальция, Са
2+
-
кальмодулина, активности МАР-киназ.
Могут регулировать процессы полимеризации актина и ло-
кализации в клетках актиновых нитей ГТФ-связывающие
белки, протеинкиназы, протеинфосфатазы, фосфоинози-
тидфосфатазы.
Показано, что актин-деполимеризующий фактор изме-
няет активность под влиянием фосфорилирования с помо-
щью Са
2+
-
зависимых протеинкиназ. У арабидопсиса был
выделен кальмодулинсвязывающий моторный вспомога-
тельный белок кинезин, причем Са
2+
-
кальмодулиновый
комплекс этого белка с кинезином ингибировал способ-
ность микротрубочек участвовать в процессах внутрикле-
точного движения. Фосфорилирование а- и |3-тубулинов ак-
тивировалось Са
2+
-
кальмодулинзависимыми и цАМФ-зави-
симыми протеинкиназами, а также протеинкиназой С
[Blume et al., 1997].
Абсцизовая кислота вызывала быструю деполимериза-
цию кортикальных актиновых нитей в замыкающих клет-
ках устьиц, причем этот процесс был опосредован входом в
клетки ионов кальция [Hwang, Lee, 2001]. Деполимеризация
медиировалась также протеинкиназами (о чем свидетельст-
вовало ее подавление стауроспорином).
Особое внимание привлекает установление факта уди-
вительного сходства трехмерной структуры цитоскелетных
вспомогательных моторных белков с G-белками. Так как
G-
белки могут непосредственно взаимодействовать с цито-
плазматическим доменом многих рецепторов в сигнальных
системах (и изменять при этом свою конформацию и актив-
ность), то этот факт позволяет допускать возможность
очень тесной связи даже между начальными звеньями сиг-
нальных систем и цитоскелетом.
Димеры тубулина имеют особенно высокое сродство с
ос-субъединицами G-белков [Крутецкая, Лонской, 1994].
Обнаружено, что активатор G-белков мастопаран вызыва-
ет (предположительно, при участии фосфоинозитольной
сигнальной системы) переход вспомогательного белка про-
филина (взаимодействующего с мономерной формой акти-
на [McCurdy et al., 2001]) из ядра в цитоплазму у клеток кор-
ней кукурузы и связанное с этим изменение структуры ак-
тиновых микрофиламентов [Baluska et al., 2001].
Выделен класс белков - пипмодулинов (связывающихся
с фосфоинозитидными фосфолипидами клеточной мембра-
ны), контролирующих освобождение фосфоинозитолбис-
фосфата (Р1Р
2
) с помощью фосфолипазы С [Lanier, Gertler,
2000] и зависящую от Р1Р
2
динамику изменения структуры и
локализации актиновых нитей, которые, в свою очередь,
определяют морфогенез клеток.
Использование ингибитора полимеризации тубулино-
вых белков оризалина, вызвавшего скопление ретикуло-
плазминовых Са
2+
-
связывающих белков вблизи плазмати-
ческой и ядерной мембран [Олиневич и др., 2001], позволи-
ло авторам сделать вывод об участии цитоскелета и ретику-
лоплазминов в преобразовании внешних сигналов.
Перестройки актиновых нитей очень чувствительны к
изменениям рН. При высоком содержании протонов акти-
новые нити более стабильны, а микротрубочки менее ста-
бильны, что не может не отразиться на состоянии цитоскеле-
та в целом [Медведев, Маркова, 1998]. Если иметь в виду,
что одним из наиболее ранних ответов клеток растений на
действие патогенов и элиситоров является подкисление ци-
тозоля, то становится очевидным, что это не может не по-
влиять на структуру и функции цитоскелета.
Описаны контролируемые цитоскелетом изменения
морфогенеза клеток, вызванные не только фитогормона-
ми, но и светом, изменением направления силы тяжести и
другими причинами. Для нас особый интерес представляют
факты действия на функционирование цитоскелета инфи-
цирования растений патогенами, приведенные в обзорной
работе [Nick, 1999]. Показано, что ядро клетки растения-хо-
зяина начинает двигаться к месту контакта с клеткой гриба
[Gross et al.,
1993]. Движение ядра осуществляется с помо-
щью актиновых микрофиламентов и обусловлено также
локальной деполимеризацией кортикальных микротрубо-
чек вокруг места контакта с грибом. Можно вызвать тор-
можение движения ядра с помощью специальных препара-
тов, причем в этом случае грибы, которые не были способ-
ны инфицировать растение, становились патогенными
[Kobayashi et al., 1997].
Авторы связывают это с ослаблени-
ем отложения каллозы на внешней поверхности плазма-
леммы, в месте контакта с патогеном.
Исследование особенностей действия элиситоров крип-
тогеина и олигогалактуронидов на цитоскелет клеток таба-
ка показало [Binet et al., 2001], что первый вызывал быстрое
и сильное разрушение сети микротрубочек, в то время как
вторые не оказывали влияния на нее. Действие криптогеи-
на положительно коррелировало с поглощением клетками
ионов кальция. В то же время имеется информация о регу-
ляции функционирования кальциевых каналов цитоскеле-
том [Thuleau et al., 1998].
Непатогенный мутант патогенного для риса гриба
Magnaporthe grisea
вызывал целый ряд защитных метаболи-
ческих реакций и перестройку актинового цитоскелета [Хи
et al., 1998].
Б
ОЛЬШОЙ
интерес вызывает информация о том, что глю-
кановые фрагменты из клеточных стенок фитофторы вы-
зывали снижение содержания мРНК одной из двух обнару-
женных изоформ тубулина, причем это было вызвано не
подавлением экспрессии гена тубулина, а деградацией тубу-
линовой мРНК, опосредованной глюканиндуцированным
повышением содержания ионов кальция в цитозоле [Ebel et
al., 2001].
Вызванная липохитоолигосахаридами (Nod-факторами)
быстрая деполимеризация актина считается главной причи-
ной успешного проникновения бактерий Rhizobium в кор-
невые волоски с последующим образованием клубеньков у
бобовых растений [Cardenas et al., 1998; Ruijter et al., 1998].
Обнаружено, что при бактериальной атаке растений одним
из наиболее быстро индуцируемых белков является цент-
рин цитоскелета [Cordeiro et al., 1998].
Итак, есть все основания считать, что цитоскелет связан
с сигнальной сетью и его изменения являются частью за-
щитного механизма против патогенных грибов и бактерий.
С другой стороны, вирусы могут использовать микротру-
бочки цитоскелета для передвижения от клетки к клетке
через плазмодесмы [Heinlein et al., 1995], и в этом случае ци-
тоскелет скорее способствует инфицированию растений, а
не их защите.
ВЗАИМОДЕЙСТВИЕ СИГНАЛЬНЫХ СИСТЕМ СО
СТРЕССОВЫМИ ФИТОГОРМОНАМИ
При изучении механизмов влияния на растения различ-
ных патогенов и элиситоров было обнаружено, что они вы-
зывают достаточно быстрое и интенсивное накопление так
называемых стрессовых фитогормонов. К ним относят со-
единения различной химической природы: абсцизовую кис-
лоту, этилен, жасмоновую кислоту (и метилжасмонат), са-
лициловую кислоту (и метилсалицилат), брассиностероиды,
системин.
Образовавшиеся стрессовые фитогормоны вызывают
синтез различных защитных соединений и повышение ус-
тойчивости как в клетках, подвергшихся действию элиси-
торов (местная или локальная устойчивость), так и на
удалении от них (системная устойчивость). Последнее
объясняется способностью большинства фитогормонов
транспортироваться на большие расстояния или вызы-
вать в клетках появление транспортных вторичных эли-
ситоров.
Стрессовый фитогормон абсцизовая кислота образуется
в результате окисления и связаного с этим разрыва углево-
дородной цепи в каротиноидах, вызванного патогенами или
элиситорами [Chernys, Zeevaart, 2000] (рис. 36). Ключевым
индуктивным ферментом, от активности которого зависит
интенсивность появления абсцизовой кислоты, является 9-
1(мс-эпоксикаротиноид-диоксигеназа.
Другой стрессовый фитогормон - летучее соединение
этилен - синтезируется в ходе последовательных реакций
(рис. 37), в которых принимают участие индуктивные фер-
менты: 1-аминоциклопропан-1-карбоксилат-синтаза (АЦКС)
и 1-аминоциклопропан-1-карбоксилат-оксидаза (АЦКО).
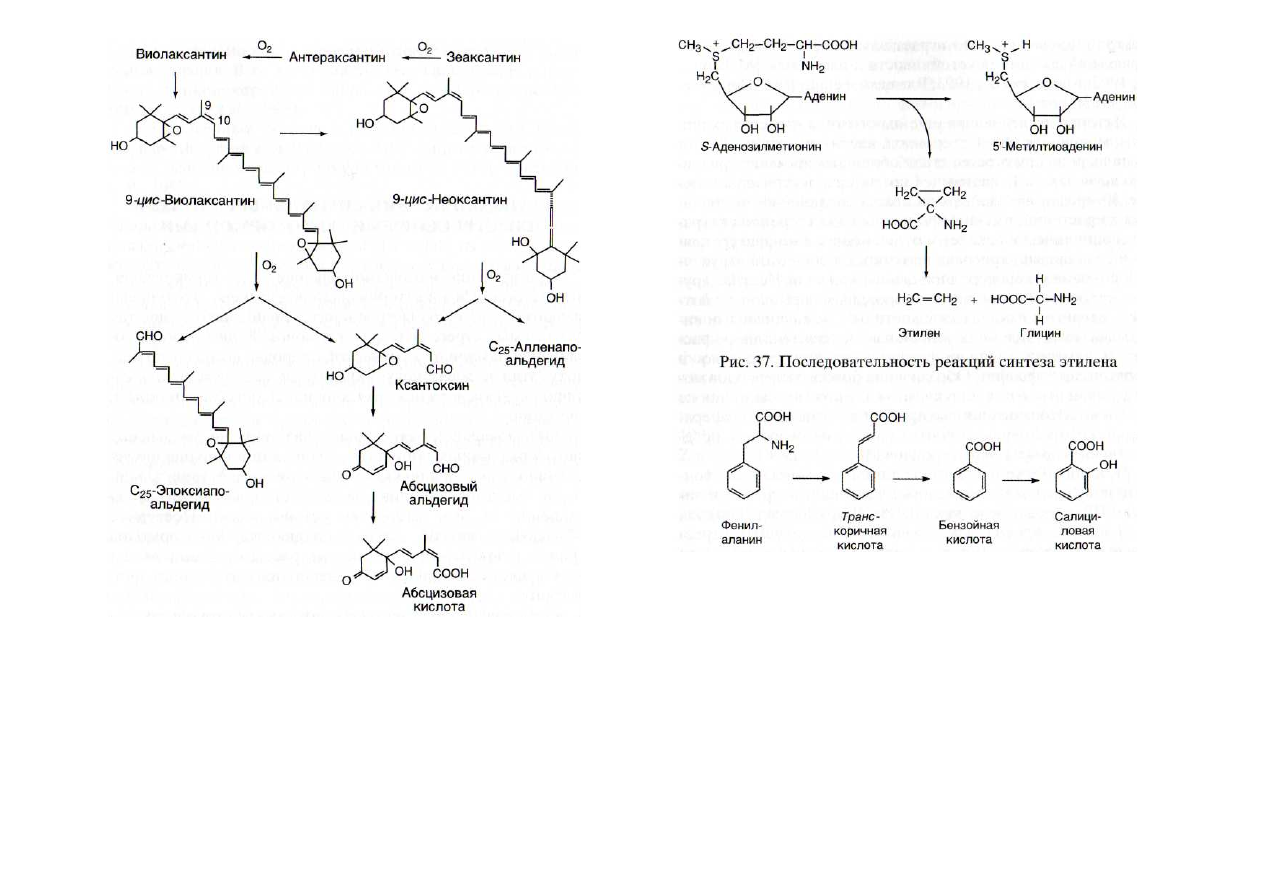
Рис. 38. Последовательность реакций синтеза салици-
ловой кислоты
Рис. 36. Последовательность реакций синтеза абсцизовой кисло-
ты [Chernys, Zeevaart, 2000]
Содержание еще одного фитогормона - салициловой
кислоты - при действии патогенов и элиситоров повышает-
ся в десятки, а в некоторых случаях более чем в сто раз
[Malamy et al., 1990; Raskin, 1992; Delaney et al., 1994; Leon et
al.,
1995]. Образуется салициловая кислота в серии реакций
из аминокислоты фенилаланина (рис. 38), причем лимити-
рует этот процесс фермент фенилаланин-аммоний-лиаза,
активируемая при действии на растения различных элиси-
торов. Фермент катализирует превращение фенилаланина
в транс-коричную кислоту, которая, по-видимому, в ре-
зультате (3-окисления образует бензойную кислоту, а пос-
ледняя с помощью бензоат-2-гидроксилазы (принадлежа-
щей к семейству цитохрома Р-450) — салициловую. Повы-
шение содержания салициловой кислоты вызывает образо-
вание из нее эфира глюкозы, откладывающегося в клеточ-
ных стенках и гидролизуемого при инфицировании расте-
ний патогенами, что вносит вклад в салицилатный
"всплеск" при патогенезе. Считается, что флоэмоподвиж-
ная салициловая кислота играет важную роль в выработке
у растений системной устойчивости к патогенам [Malamy et
al., 1993; Metraux et al., 1993; Rasmussen et al.,
1991; Запроме-
тов, 1993; и др.].
История исследования еще одного типа стрессовых фи-
тогормонов - брассиностероидов, насчитывает уже 20 лет,
но на их роль при стрессе стали обращать внимание сравни-
тельно недавно. В настоящее время охарактеризовано бо-
лее 40 представителей этого класса соединений, выделен-
ных из растений, имеющих типичный для стероидов тетра-
гетероциклический скелет и отличающихся конфигурация-
ми и радикалами, присоединеными к циклической структу-
ре и боковой углеводородной цепи. Так же как в случае дру-
гих стеролов (в том числе стероидных гормонов живот-
ных), их синтез начинается с ацетил-КоА, а основными ин-
термедиатами являются мевалонат, изопентенилпирофос-
фат и диметилаллилпирофосфат, геранилпирофосфат и
фарнезилпирофосфат. Образование брассиностероидов мо-
жет усиливаться под влиянием патогенов, но было также
отмечено и торможение экспрессии гена метилтрансфера-
зы, от которой зависел синтез одного из основных пред-
шественников брассиностероидов [Shi et al., 1996].
При обработке растений экзогенными брассиностерои-
дами повышается устойчивость к патогенным грибам и ви-
русам [Шакирова, Безрукова, 1998; Khripach et al., 2000].
Выше уже отмечалось, что патогениндуцируемые стрес-
совые фитогормоны могут вызывать ответную реакцию на
достаточно большом удалении от места инфицирования
ткани - системный иммунитет растений, проявляющийся в
устойчивости к самым различным патогенам.
Внимание исследователей механизма формирования си-
стемного иммунитета привлек сравнительно небольшой по-
липептид, состоящий из 18 аминокислотных остатков, на-
званный системином. Ряд авторов признали его первым
идентифицированным фитогормоном полипептидной при-
роды [McGurl et al., 1992; Slosarek et al., 1995; Bergey et al., 1996;
Bowles, 1998; Constabel et al., 1998; Ryan, Pearce, 1998; Chao et
al., 1999; Dombrowski et al., 1999; Sheer, Ryan,
1999]. Интен-
сивное образование системина, который сначала считали
раневым сигналом, происходит при механическом повреж-
дении тканей растений (например, травоядными животны-
ми или листогрызущими насекомыми), действии на расте-
ния различных патогенов, элиситоров, обезвоживания, за-
соления и других стрессоров. В связи с этим системин мо-
жет считаться стрессовым фитогормоном. Этот пептид яв-
ляется продуктом частичной деградации (посттрансляцион-
ной модификации) более крупного предшественника - про-
системина, состоящего из 200 аминокислот. Замещение в
системине остатка пролина-13 и тирозина-17 аланином при-
водит практически к полной потере элиситорной активно-
сти системина [Pearce et al., 1993]. Накопление системина
носит двухфазный характер, что объясняется его разруше-
нием за счет протеолиза связи по местоположению лизи-
на-14. При N-метилировании этой связи системин становит-
ся более устойчивым к действию протеаз, и активность сис-
темина становится более продолжительной, по сравнению с
контрольным вариантом [Schaller, 1998]. Исследования кон-
формации системина привели к неоднозначным результа-
там. С одной стороны, на основании данных ЯМР-спектро-
скопии сделан вывод об отсутствии в этом полипептиде
жесткой структуры типа спирали или ленты [McGurl, Ryan,
1992], с другой [Slosarek et al., 1995] - предложена модель Z-
подобной ленточной (3-структуры системина.
В интактных растениях ген просистемина может быть
отнесен к числу спящих. Он состоит [McGurl et al., 1992] из
десяти интронов и одиннадцати экзонов, десять из которых
организованы в пять гомологичных пар. Данные о роли
системина в качестве фактора индукции системной устой-
чивости растений подкрепляются тем, что это — флоэмомо-
бильный полипептид [Schaller, Ryan, 1996].
Фитогормон может опосредовать лишь часть ответа
клеток на тот или иной элиситор. Обнаружено, например,
что метилжасмонат индуцирует накопление сесквитерпен-
циклазы (одного из ключевых ферментов синтеза терпено-
идных фитоалексинов) лишь в следовых количествах, в от-
личие от быстрого и интенсивного образования этого фер-
мента в культуре клеток табака под влиянием грибных эли-
ситоров [Mandujano-Chavez et al., 2000].
Один из стрессовых фитогормонов - жасмоновая кисло-
та, а также метилжасмонат образуются (см. рис. 19) в ходе
реакций элиситориндуцируемого липоксигеназного метабо-
лизма [Vick, Zimmerman, 1987; Гречкин, Тарчевский, 1999].