ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 27.11.2019
Просмотров: 4038
Скачиваний: 98

Оказалось, что элиситоры могут также активировать фер-
менты синтеза этилена [Spanu et al., 1991; Bowler, Chua,
1994; Oetiker et al.,
1997], причем повышенные концентра-
ции ионов кальция усиливали этот процесс [Gallardo et al.,
1999].
Элиситоры индуцировали экспрессию некоторых генов,
кодирующих ферменты синтеза абсцизовой кислоты
[Chernys, Zeevaart,
2000]. Наблюдалось также повышение
содержания салициловой кислоты [Malamy et al., 1990;
Klessig, Malamy,
1994] и системина [Bergey et al., 1996], уча-
ствующих в индукции системного ответа растений на дейст-
вие того или иного патогена. Некоторые исследователи
считают таким индуктором защитных реакций растений пе-
рекись водорода, которая может выступать в этой роли не-
зависимо от салицилата. Было, например, обнаружено, что
у растений табака сублетальные концентрации перекиси
водорода вызывали системную индукцию образования ос-
новных и кислых защитных белков (PR) и повышение ус-
тойчивости растений табака к патогенам, но этот эффект
был быстрее и сильнее, когда сопровождался некротиче-
скими изменениями в листьях [Chamnongpol et al., 1998]. Ос-
тается неясным, был ли системный ответ следствием непо-
средственного действия перекиси водорода или был опосре-
дован другими сигнальными молекулами. Ими могут быть
интермедиаты или продукты функционирования различных
сигнальных систем клеток. Обнаружено, что жасмонат и
метилжасмонат активируют липоксигеназную [Bell, Mullet,
1991; Melan et al., 1993; Avdiushko et al., 1995; Jensen et al.,
1997; Voros et al.,
1998] и НАДФН-оксидазную [Tamagami et
al.,
1997] сигнальные системы. Этилен активировал липок-
сигеназную и МАР-киназную [Voeste, Kieber, 1998; Iten et al.,
1999] системы. Абсцизовая кислота ингибировала адени-
латциклазную [Корчуганова и др., 1998], но активировала
липоксигеназную [Melan et al., 1993] (ингибируя в то же вре-
мя превращение фитодиеновой кислоты в жасмоновую
[Laudert, Weiler,
1998]) и кальциевую [Mikami et al., 1998;
Owen, 1988; Staxen,
1999], МАР-киназную [Knetsch et al.,
1996; Iten et al.,
1999], НАДФН-оксидазную [Guan,
Scandalios,
1998] и фосфатидатную [Munnik et al., 1995;
Ritchie, Gilroy, 1998; Munnik,
2001] сигнальные системы.
Абсцизовая кислота вызывала накопление одного из ин-
термедиатов NO-сигнальной системы - цАДФрибозы [Yu et
al., 1997; Leckie et al., 1998; Walden, 1998].
Салицилат активировал липоксигеназную [Feussner et
al., 1997b],
МАР-киназную [Zhang, Klessig, 1997; Iten et al.,
1999; Romeis et al.,
1999], НАДФН-оксидазную [Chen et al.,
1993] и NO-синтазную [Klepper, 1991; Van Camp et al., 1998];
системин-кальциевую [Bergey, Ryan, 1999], липоксигеназ-
ную [Constabel et al., 1995; Narvaez-Vasquez et al., 1999;
Orozco-Cardenas, Ryan,
1999], НАДФН-оксидазную [Stennis
et al.,
1998], МАР-киназную [Stratmann, Ryan, 1997; Meindl et
al.,
1998] и протонную [Schaller, Oecking, 1999], а перекись
водорода - липоксигеназную [Macri et al., 1994] и НАДФН-
оксидазную [Leon et al., 1995; Dorey et al., 1999] сигнальные
системы.
Обнаружено, что экзогенный системин вызывает акти-
вацию липоксигеназной [Narvaez-Vasquez et al., 1999; Sheer,
Ryan,
1999], кальциевой [Bergey, Ryan, 1999] и MAPK-
[ Stratmann, Ryan,
1997] сигнальных систем.
Стрессовые фитогормоны могут усиливать или затор-
маживать образование друг друга. Так, оказалось, что жас-
монат вызывает значительное снижение содержания в
клетках абсцизовой кислоты [Hays et al., 1999], а салицилат
ингибирует синтез жасмоната [Репа-Cortes et al., 1993; Doares
et al., 1995a]
и этилена [Leslie, Romani, 1986]. Последнее объ-
ясняется тем, что салициловая кислота - это ингибитор не
только каталазы, но и ряда других железосодержащих фер-
ментов, в том числе ключевого фермента синтеза этилена -
1-
аминоциклопропан-1-карбоксилат-оксидазы [Chen et al.,
1997]. Этилен и метилжасмонат синергично индуцировали
синтез защитных белков [Xu et al., 1994]. Салицилат подав-
лял жасмонатиндуцированное [Репа-Cortes et al., 1993;
Doares et al., 1995a],
АБК- [Taipalensuu et al., 1997] и систе-
мининдуцированное [Doares et al., 1995a] образование бел-
ков, а абсцизовая кислота затормаживала этилениндуциро-
ванный синтез клетками растений глюканаз, но усиливала
такой синтез хитиназ [Rezzonico et al., 1998]. Метилжасмо-
нат индуцировал синтез рецептора системина, локализован-
ного в плазмалемме и, вследствие этого, стимулировал появ-
ление вызванных системином ответных реакций [Sheer,
Ryan,
1999], а системин индуцировал синтез абсцизовой
кислоты [Chao et al., 1999].
Салицилат приводил к экспрессии стероидной сульфо-
трансферазы, вызывающей подавление брассиностероидной
активности [Rouleau et al., 1999]. В то же время синтез стеро-
идной гидроксилазы семейства цитохромов Р-450, играющей
центральную роль в синтезе брассинолида, не изменялся под
влиянием экзогенных салицилата, этилена и жасмоната, но
авторегулировался брассинолидом [Mathur et al., 1998].
Использование мутантов растений позволило установить,
что жасмонат и этилен "включают" одни сигнальные пути, а
салицилат - другие [Dong, 1998], однако были также установ-
лены различия в особенностях протекания ответных реак-
ций, вызванных жасмонатом и этиленом [Penninckx et al.,
1998]. На растениях арабидопсиса было показано, что сали-
цилат и жасмонат вызывают аддитивный защитный ответ на
инфицирование патогенами, но не в тех случаях, когда иссле-
довались мутанты с заблокированным салицилатным или
жасмонатным сигнальными путями [Van Wees et al., 2000].
Авторы считают, что эти пути являются комплементарными
и нет оснований говорить об их пересечении (cross-talk) и пе-
ретоке сигналов из одного в другой.
О степени автономности или взаимовлияния сигнальных
путей, "включаемых" различными стрессовыми фитогор-
монами, можно судить по индуцированию ими образования
различных транскриптов и синтеза различных белков. Так,
у листьев редиса под влиянием метилжасмоната и этилена
активировалась экспрессия генов дефенсинов [Terras et al.,
1998; Shah et al.,
1999], но не защитных белков PR1, а сали-
циловая кислота вызывала активацию локального синтеза
PR1,
но не системного синтеза дефенсинов. В то же время
метилжасмонат не действовал на синтез некоторых изо-
форм дефенсинов [Epple et al., 1997]. Как правило, салици-
ловая кислота не индуцировала образование дефенсинов
[Epple et al., 1997; Terras et al., 1998; Shah et al., 1999].
У томатов салицилат ингибировал, а абсцизовая кисло-
та и системин активировали синтез мРНК леициновых ами-
нопептидаз, вызванный повреждением тканей, причем два
последних фитогормона действовали синергично. В то же
время синтез ингибиторов протеиназ не зависел от экзоген-
ной абсцизовой кислоты [Chao et al., 1999].
К сожалению, упомянутые исследования, в которых
проводилось сравнительное изучение влияния различных
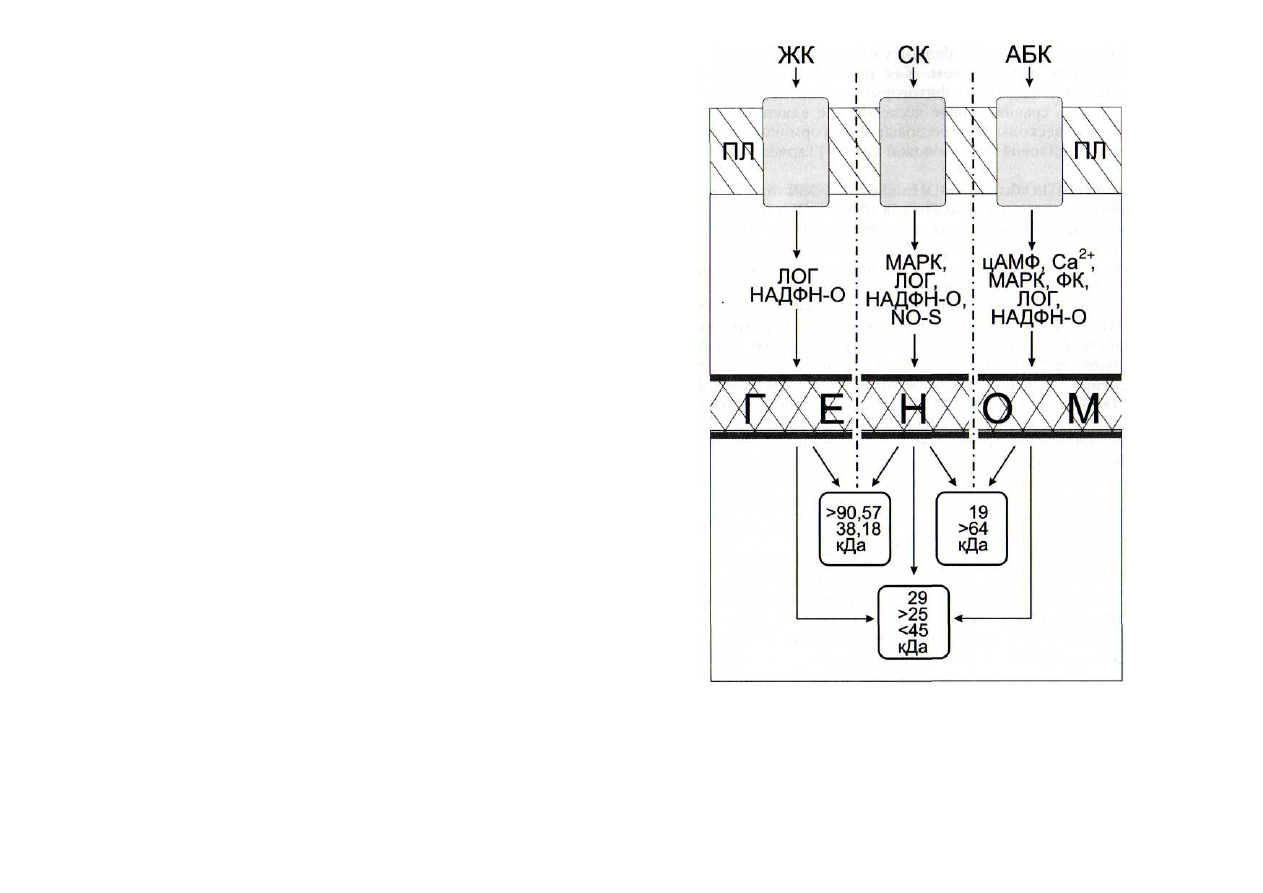
Рис. 39. Схема влияния абсцизовой (АБК), салициловой (СК) и
жасмоновой (ЖК) кислот на синтез полипептидов [Тарчевский и
др., 2001]
ЛОГ - липоксигеназная сигнальная система; МАРК - МАР-киназ-
ная сигнальная система; НАДФН-О - НАДФН-оксидазная сигнальная
система; ПЛ - плазмалемма; ФК - фосфатидатная сигнальная система;
цАМФ - аденилатциклазная сигнальная система; Са
2+
-
кальциевая сиг-
нальная система; NO-S - NO-синтазная сигнальная система
фитогормонов на синтез белков у одного и того же объек-
та, являются исключением. Как правило, анализируется
действие только одного фитогормона. Это побудило нас
предпринять сравнительное исследование влияния на син-
тез белков нескольких стрессовых фитогормонов - абсци-
зовой, салициловой и жасмоновой кислот [Тарчевский и др.,
2001].
Экзогенные абсцизовая и салициловая кислоты индуци-
ровали образование новых белков 19 и 29 кДа у проростков
гороха (рис. 39). Экзогенный жасмонат вызывал появление
двух новых полипептидов 29 и 96 кДа и исчезновение поли-
пептида 104 кДа. Из результатов проведенных исследова-
ний следует, что образование нового полипептида 29 кДа,
интенсивно метящегося и не проявляющегося на радиоавто-
графах гелей контрольного варианта, - характерный ответ
на действие всех исследовавшихся нами стрессовых фито-
гормонов, так же как усиление интенсивности синтеза по-
липептида 25 кДа и торможение синтеза полипептида
45 кДа [Тарчевский и др., 2001]. Это, по-видимому, можно
рассматривать как неспецифический ответ растений. Были
обнаружены и различия в наборе образующихся полипеп-
тидов и интенсивности их синтеза. Так, при действии жасмо-
новой кислоты наблюдалось появление полипептида
96 кДа, исчезновение белка 104 кДа и усиление образования
полипептидов 35 и 71 кДа, чего не наблюдалось при обра-
ботке растений другими фитогормонами.
Сопоставляя данные литературы с полученными нами
результатами, необходимо обратить внимание на то, что
две важнейшие сигнальные системы (липоксигеназная и
НАДФ-оксидазная) "включаются" каждым из исследовав-
шихся нами стрессовых фитогормонов. По всей вероятно-
сти, это и определяет неспецифичность ответа со стороны
части генома клеток (образование нового полипептида
29 кДа, усиление синтеза полипептида 25 кДа и торможение
синтеза полипептида 45 к Да). Специфичность экспрессии
генов и вследствие этого включения [
14
С]-лейцина в поли-
пептиды может объясняться своеобразием индукции от-
дельными фитогормонами различных сигнальных систем
клеток.
РЕГУЛЯЦИЯ ИОННЫХ ПОТОКОВ
ИНТЕРМЕДИАТАМИ
СИГНАЛЬНЫХ СИСТЕМ
Тот факт, что блокаторы ионных каналов ингибировали
синтез фитоалексинов [Ward et al., 1995; Roos et al., 1998],
свидетельствует об участии ионов в работе сигнальных сис-
тем клеток и формировании ответа на инфицирование рас-
тений патогенами. Это положение подтверждается также
фактами "включения" синтеза фитоалексинов (даже при
отсутствии элиситоров) целым рядом ионофоров, способ-
ных переносить в клетку из окружающей среды протоны
или ионы кальция, а также отсутствием действия элисито-
ров при "декальцинировании" или "депротонизации" среды.
Для исследования роли ионных потоков в сигнальном мета-
болизме клеток применяют такие ионофоры, как 2,4-динит-
рофенол (протонофор), валиномицин (специфический К
+
-
переносчик), нигерицин (К
+
/Н
+
-
обменник), моненсин
(Ка
+
/Н
+
-
обменник), А23187 (специфический Са
2+
-
перенос-
чик), иономицин (специфический Са
2+
-
переносчик); канало-
формеры грамицидины А и Д (образуют каналы, через ко-
торые проходят Н
+
, К
+
), полиеновые антибиотики амфоте-
рицин и нистатин (образуют поры, через которые проходят
катионы и анионы); блокаторы ионных каналов тетраэтил-
аммоний (блокирует К
+
-
каналы), верапамил и нифедипин
(блокируют Са
2+
-
каналы), лантаниды (блокируют прак-
тически все каналы; но чаще всего используются для блока-
ды Са
2+
-
каналов), этакриновая кислота (блокатор анион-
ных каналов). Используются также ингибиторы (эритро-
зин В, диэтилстилбестрол, ванадат) и активаторы (фузикок-
цин) Н
+
-
АТФаз.
Очень важно, что изменения концентраций ионов и свя-
занной с ними трансмембранной разности потенциалов -
преходящи, так же как многие более поздние метаболиче-
ские ответы клеток на действие элиситоров. Подавление со
временем возбуждения клетки, индуцированного внешним
сигналом, является одним из основных принципов регуля-
ции биологических систем. Выяснение механизмов или "ры-
чагов" такой супрессии - одна из очень важных задач фи-
зиологии растений. Можно предположить, что если ранняя
активация сигнальных систем клеток зависит от трансмем-
бранного изменения концентраций определенных ионов, то
они, в свою очередь, могут испытывать на себе регулирую-
щее действие со стороны сигнальных систем.
Действительно, происходит активация ионных каналов
относительно небольшими концентрациями интермедиа-
тов сигнальных систем. Показана активация кальциевых
каналов интермедиатами сигнальных систем: аденилатцик-
лазной (цАМФ), кальциевой (ИФ
3
и ИФ
4
, Са
2+
), NO-
син-
тазной (цАДФРиб и цГМФ) [Авдонин, Ткачук, 1994]. От-
мечено также ингибирование кальциевых каналов интер-
медиатами липоксигеназной системы (полиеновыми жир-
ными кислотами и их гидропероксипроизводными) [Авдо-
нин, Ткачук, 1994], а также повышенными концентрация-
ми ионов кальция (случай автокаталитического подавле-
ния нарастания ионов кальция в цитоплазме).
Все большее автокаталитическое повышение концент-
раций указанных вторичных посредников приводит к акти-
вации кальциевых насосов клетки, выводящих эти ионы из
цитозоля (см. рис. 13) и, таким образом, снижающих их ак-
тивирующее влияние на сигнальные системы. Стимуляция
кальциевых и протонных помп вызывает реполяризацию
мембран (плазмалеммы и тонопласта). К такому же эф-
фекту должны приводить активация калиевых каналов (на-
пример, повышающимися концентрациями цАМФ) и усили-
вающийся выход СГ. Повышают активность Н
+
-
АТФаз
плазмалеммы ионы кальция, лизофосфолипиды [Li et al.,
1994а; Scherer, 1996a,b], полиеновые жирные кислоты
[Scherer,
1996], цАМФ и цГМФ, причем два последних сиг-
нальных интермедиата могут прямо, минуя стадию актива-
ции протеинкиназ, связываться с белками ионных каналов
(цАМФ- и цГМФ-зависимых каналов) [Li et al., 1994a; Дячок
и др., 1997]. Имеются также противоположные данные о
влиянии на активность мембранных Н
+
-
АТФаз [Владими-
ров, 1998] низких и высоких, но физиологических концент-
раций ионов кальция.
Промежуточные продукты различных сигнальных сис-
тем могут оказывать влияние на функционирование ион-
ных каналов и помп непосредственно или с помощью соот-
ветствующих протеинкиназ [Conrath et al., 1991; Li et al.,
1994b].
Кальциевые каналы активируются такими сигналь-
ными интермедиатами, как ИФ
3
, ИФ
4
[Gilroy et al., 1990,
1993; Trewavas,
1999; и др.], цАМФ [Авдонин, Ткачук, 1994;
Дячок и др.,1997; Volotovsky et al., 1998], цАДФ-рибоза,
цГМФ [Авдонин, Ткачук, 1994; Volotovsky et al., 1998]. Са
2+
-
АТФазы активируются Са
2+
-
кальмодулином, но ингибиру-
ются гидропероксипроизводными полиеновых жирных кис-
лот [Владимиров, 1998]. Активируют [Scherer, 1996a,b; Ши-
шова, 1999] (или ингибируют [Шишова, 1999]) Н
+
-
АТФазу
плазмалеммы повышенные концентрации ионов кальция
(Kinoshita et al., 1995; Scherer, 1996a,b],
Са
2+
-
зависимые про-
теинкиназы [Schaller, Oeckingb, 1999], лизофосфатиды, по-
лиеновые жирные кислоты [Scherer, 1996a,b], цАМФ и
цГМФ [Palmgren, 1991; Vera-Estrella et al., 1994]. Выход
ионов калия из клеток усиливается под влиянием цАМФ,
активирующего протеинкиназы и с их помощью - калие-
вые каналы плазмалеммы [Li et al., 1994a; Tang, Hoshi, 1999].
Следующая за изменением ионных потоков местная де-
поляризация плазмалеммы, вызванная разрушением клеток
листогрызущими насекомыми и другими способами меха-
нического повреждения, приводит к распространению элек-
трического импульса, которое захватывает соседние клет-
ки и проводящую систему растений и может участвовать в
индукции системного иммунитета в удаленных от места пов-
реждения участках растений [Stankovic , Davies, 1996].
Деполяризация ПЛ представляет собой сдвиг мембран-
ного потенциала покоя от (-140...-200 мВ) до более поло-
жительных значений под влиянием различных воздействий,
в том числе элиситорных сигналов. Существует несколько
механизмов, от функционирования которых зависит депо-
ляризация плазмалеммы и тонопласта: активация кальцие-
вых каналов, анионных каналов, ингибирование протонной
АТФазы и др. Имеются потенциалзависимые Са
2+
-
каналы,
потенциалзависимые К
+
-
каналы, анионные каналы с отно-
сительно малым (секунды) или более продолжительным
(минуты) временем деполяризации. Местное повышение
концентрации ионов кальция, а также вызванное этим сни-
жение величины трансмембранного электрического потен-
циала могут привести к открыванию калиевых и анионных
(хлорных) каналов [Trewavas, 1999] и транспорту этих ионов
из цитоплазмы за пределы клетки или в тонопласт.
Очень важными мишенями действия цАМФ у живот-
ных, помимо протеинкиназ А, являются циклонуклеотидре-
гулируемые ионные каналы. Существуют доказательства
того, что и у растений цАМФ может регулировать К
+
-
кана-
лы [Li et al., 1994a; Bolwell, 1995; Kurosaki, 1997], Са
2+
-
кана-
лы [Kurosaki et al., 1994; Volotovsky, 1998; Leng et ah, 1999],
Cl-
каналы [Gabriel et al., 1999], Н
+
-
каналы [Bolwell, 1999]
мембран клеток, возможно, минуя их фосфорилирование с
помощью протеинкиназ [Bolwell, 1995; Walden,1998]. Более
того, клонированные белки ионных каналов растений име-
ли циклонуклеотидсвязывающие места [Bolwell, 1995; Leng
et al.,
1999]. Наличие в белках ионных каналов как кальмо-
дулинсвязывающего, так и цАМФ-связывающего доменов
позволяет сделать вывод о совместном влиянии аденилат-
циклазной и кальциевой сигнальных систем на ионные по-
токи в клетках [Arazi et al., 2000].
ВЗАИМОДЕЙСТВИЕ СИГНАЛЬНЫХ СИСТЕМ
Различные сигнальные системы могут включаться не
только разными элиситорами, но даже одним элиситором.
Так, криптогеин, секретируемый Phytophthora crypto-
gea,
связывается с рецептором (рецепторами) в плазмати-
ческой мембране и индуцирует активацию нескольких
сигнальных систем (МАР-киназной, кальциевой и супер-
оксидсинтазной) [Allan, Fluhr, 1997; Lebrun-Garcia et al.,
1999]. Обнаружена также активация криптогеином липок-
сигеназной сигнальной системы [Rusterucci et al., 1999].
У гликопротеидного элиситора патогенного гриба
Verticillium
белковая часть обеспечивала сигнальные пу-
ти, ведущие к синтезу фитоалексинов, а углеводная инду-
цировала окислительный взрыв, зависящий главным об-
разом от функционирования НАДФН-оксидазной сиг-
нальной системы. Элиситор, продуцируемый Pseudomonas
syringae,
взаимодействует с рецепторной киназой, которая
активирует несколько (по крайней мере три) факторов
регуляции транскрипции [Zhou et al., 1997], что обеспечи-
вает дивергенцию сигнального потока. Возможность по-
добной дивергенции поддерживается и другими исследо-
вателями (рис. 40), учитывающими факты множествен-
ности мест фосфорилирования у киназ и олигомеризации
рецепторных киназ, увеличивающей число мест фосфо-
рилирования, по сравнению с мономерной неактивной
формой [Cohen, 2000].
Высказываются мнения о возможности как раздельного
(параллельного) функционирования сигнальных систем
[Doares et al., 1995a; Desikan et al., 1996; Bolwell et al., 1998;
Heo et al.,
1999]: липоксигеназной и НАДФН-оксидазной
[Doares et al., 1995b],
кальциевой и НАДФН-оксидазной
[Heo et al., 1999], NO-
синтазной и НАДФН-оксидазной