ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 09.11.2023
Просмотров: 648
Скачиваний: 2
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
СОДЕРЖАНИЕ
Лимфоидная ткань, ассоциированная со слизистыми оболочками
Т-лимфоциты и их субпопуляции у детей
В-лимфоциты и иммуноглобулины у детей
Аутовоспалительные заболевания (периодические лихорадки)
ГЛАВА 5 АЛЛЕРГИЧЕСКИЕ ЗАБОЛЕВАНИЯ
Патогенез и клиническая патофизиология
Диагностические критерии аллергических заболеваний
Выявление факторов риска (триггеров) и уменьшение их воздействия
Элиминационные мероприятия по уменьшению кон- такта с пыльцой:
Элиминационные мероприятия по уменьшению кон- такта с аллергенами грибов:
ОСНОВЫ КЛИНИЧЕСКОЙ ИММУНОЛОГИИ И АЛЛЕРГОЛОГИИ ДЕТСКОГО ВОЗРАСТА
ри и ребенка при беременности, в том числе лимфоцитами. Примерно 5% лимфоцитов новорожденного – материнские. Вероятная польза данного процесса для ребенка может состоять в том, что зрелые иммунокомпетентные клетки матери способствуют созреванию и «обучению» иммунной системы. Польза фетально- го микрохимеризма для матери может состоять в том, что клетки ребенка, проникшие в материнский организм, способствуют под- держанию толерантности матери к антигенам плода и сохранению беременности. Возможен и вред микрохимеризма как для ребенка, так и для матери. Многолетнее носительство клеток с чужим гено- типом может индуцировать аутоиммунную патологию и неопла- зии. Первичные иммунодефицитные состояния сопровождаются активным материнским микрохимеризмом, являющимся одним из механизмов, объясняющих аутоиммунные расстройства при первичных иммунодефицитах, развитие реакции трансплантант против хозяина (проявляется эритематозной сыпью).
- 1 ... 14 15 16 17 18 19 20 21 ... 40
Т-лимфоциты и их субпопуляции
Т-лимфоциты – основной участник так называемой клеточной формы иммунного реагирования. В отличие от В-лимфоцитов, функция которых реализуется через гуморальные продукты – антитела, Т-лимфоциты разрушают чужеродные клетки (транс- плантаты, опухолевые клетки, вирус-трансформированные клетки) при непосредственном контакте с данными клеточны- ми формами. Распознавание чужеродных клеток осуществляется антиген-распознающими Т-клеточными рецепторами, экспрес- сирующимися на поверхности Т-лимфоцитов. Кроме цитоток- сической функции Т-лимфоциты осуществляют регуляторное воздействие на гуморальный и клеточный иммунный ответ по- средством продукции цитокинов.
Самый начальный этап дифференцировки Т-лимфоцитов осу- ществляется в костном мозге, когда из стволовой кроветворной клетки (СКК) образуется наиболее ранний предшественник, мигрирующий в тимус. Именно в нем происходят основные со- бытия клеточной дифференцировки, приводящие к формирова- нию субпопуляций Т-клеток: CD8+ Т-лимфоцитов (цитотоксиче- ских – Тс) и CD4+ Т-лимфоцитов (Т-хелперов, Th). Разделение Т-лимфоцитов на CD4+ и CD8+ имеет место только в тимусе, в ходе дифференцировки, но не на периферии. В норме соотноше- ние Тh/Тc составляет 2:1. При этом Т-хелперы выходят из тиму-
са не активированными клетками (“наивными” или “нулевыми” Th0) и в ходе иммунного ответа превращаются в Т-хелперы 1-ого и 2-ого типа (Th1 и Th2), опосредующие реакции клеточного им- мунного ответа (Th1), протекающего по механизму хронического воспаления (“реакции гиперчувствительности
замедленного ти- па”), либо реакции гуморального иммунного ответа, связанного с выработкой антител (Th2).
Между собой Тh1 и Th2 различаются по набору секретируемых цитокинов. Главными цитокинами Th1 являются провоспали- тельные молекулы ИФН-γ, ФНО-α, ИЛ-2, главными цитокинами Тh2 – ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-10, а также трансформирующий фактор роста β (ТФРβ). Действие ИЛ-10 и ТФРβ носит противо- воспалительную направленность.
При активации «наивная» СD4+(Th0)-клетка дифференциру- ется либо в сторону Тh1, либо Th2. В генетически здоровом ор- ганизме отклонение иммунного ответа в сторону Тh1 или Th2 зависит от внешних факторов: химических свойств антигена, его дозы, пути попадания во внутреннюю среду, типа адьювантов или инфекций, кратности иммунизации и др. По современным пред- ставлениям выбор пути специализации Th0 в большей степени зависит от медиаторного фона, возникающего на участке презен- тации антигена. Если презентация антигена происходит в присут- ствии ИЛ-12, ИЛ-18, ИЛ-21 и(или) ИФН-γ, то Th0 превращаются в Тh1, который в свою очередь продуцирует ИФН-γ, ИЛ-2, ФНОα, ФНОβ, а также ИЛ-3, гранулоцитарно-макрофагальный колоние- стимулирующий фактор (ГМ-КСФ) и другие цитокины. При этом источниками ИЛ-12, ИЛ-18 и ИЛ-21 выступают АПК, а источни- ком ИФН-γ могут быть естественные киллеры (NK-клетки).
В случае презентации антигена через молекулу главного ком-
плекса гистосовместимости II класса в присутствии ИЛ-4 Th0 пе- реходят в Th2, которые секретируют ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-10, а также ИЛ-3, ГМ-КСФ и некоторые другие медиаторы. Источ- никами ИЛ-4 могут быть тучные клетки, тромбоциты, активи- рованные ранее Th2. Участие тучных клеток в этом процессе не
случайно, так как проникновение антигена во внутреннюю среду организма, где расположены эти клетки, всегда сопровождается их дегрануляцией и секрецией цитокинов.
Несмотря на антагонистические отношения, и Тh1, и Th2 секре- тируют ИЛ-3 и ГМ-КСФ. Интерлейкин-3 влияет на СКК и стиму- лирует образование новых клеток лимфоидного ряда (как Т-, так и
50
Таблица 10
Характеристика популяций и субпопуляций Т-лимфоцитов
| Характе- ристики | Т-цито- токсиче- ские | Тh0 | Тh1 | Тh2 | Treg | Th17 | NKT |
| Происхож- дение | Из клеток предше- ственников в тимусе | Из клеток предше- ственников в тимусе | Из Тh0, переход Тh0→Тh1 в ходе иммунного ответа в периферических лимфоидных органах | Из Тh0, переход Тh0→Тh2 в ходе иммунного ответа в периферических лим- фоидных органах | Из Т-кле- ток, пре- имуще- ственно в тимусе. | Из активиро- ванных CD4+ лимфоцитов | Из клеток пред- шественников в тимусе |
| Основные секрети- руемые медиаторы | ИЛ-2, ФНОα, ИФН-γ | ИЛ-2, ИЛ-4, ИФН-γ | ИФН-γ, ФНОα, ИЛ-2 | ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-10 | ИЛ-10, ТФРβ | ИЛ-17, ИЛ-22, ИЛ-21 | ИФН-γ, ИЛ-2, ИЛ-10, ИЛ-17, ФНОα |
| Поверх- ностный маркер | СD8+ | СD4+ | СD4+ | СD4+ | CD4+ CD25+ | CD4+ | CD4+ CD8+ CD16+ CD56+ |
| Роль в иммунном ответе | Уничтоже- ние вирус- инфици- рованных клеток | Первичное распознава- ние антигена и превраще- ние в Тh1 или Th2 | Клеточный имму- нитет, стимуляция макрофагов при хроническом вос- палении (реакция ГЗТ), торможение функций Тh2 | Стимуляция В-лимфо- цитов к превращению в плазматические клетки и секреция антител (гуморальный иммуни- тет), атопия, торможе- ние функций Тh1 | Супрес- сорная функция в отноше- нии Th1 и Th2 | Антимикроб- ная защита эпителиальных и слизистых барьеров, участие в противоопухо- левом иммунном ответе | Клетки врожден- ного иммунитета, цитотоксическая и регуляторная функция, иммунный надзор |
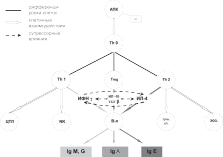
Рис. 5. Th1/Th2 - парадигма
Примечания: АПК – антиген-презентирующая клетка; АГ – антиген; Th
-
T-хелперы; Treg – T-регуляторные лимфоциты, ЦТЛ – цитотоксические лимфоциты; В-л. – В-лимфоциты; NK – NK-клетки; тучн. кл. – тучные клетки; эоз. – эозинофилы.
В-лимфоцитов). В свою очередь ГМ-КСФ, действуя на СКК, вызыва- ет образование и активацию новых клеток моноцитарно-макрофа- гального ряда и гранулоцитов. При завершении иммунного ответа по варианту гиперчувствительности замедленного типа (ГЗТ) раз- вивается хроническое воспаление. Макрофаги при этом выступают в роли эффекторных клеток. При завершении иммунного ответа по гуморальному пути макрофаги и гранулоциты (главным образом, нейтрофилы) выступают в роли клеток-дворников и уничтожают разрушенные микробы и структуры тканевого дендрита.
Помимо Тh1 и Th2 существует еще одна субпопуляция CD4+ Т-лимфоцитов – естественные Т-регуляторные клетки (Тreg, СD4+CD25+Foxp3), или Т-хелперы третьего типа (Th3), кото- рые обеспечивают главным образом синтез противовоспали- тельных цитокинов ТФРβ и ИЛ-10, подавляющих иммунный ответ, а также синтез IgA и поддержание иммунологической толерантности. Различия субпопуляций Т-лимфоцитов пред- ставлены на рис. 5 и в табл. 10.
Различные варианты иммунного реагирования в результате кооперации лимфоцитов представлены в