ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 04.12.2023
Просмотров: 760
Скачиваний: 1
СОДЕРЖАНИЕ
Сабақ 2 Жасушаның генетикалық аппараты
Сабақ 3 . Тақырып: Сомалық және жыныс жасушалардың бӛлінуі. Тақырыптың негізгі сұрақтары:
Постмитоздық кезең (пресинтездік) кезең G1:
Гаметогенез (оогенез, сперматогенез), сатылар сипаттмамасы. Мейоз анықтамасы және
Митоз және мейоздың медициналық маңызы.
2 сурет. Көз бұршағының индукциясы.
Онтогенездің жасушалық механизмдерi.
Даму ақауларының сызбасы ДТБА жіктелуі:
Сабақ 5 АҚПАРАТТЫҚ – ДИДАКТИКАЛЫҚ БЛОК
Аутосомды-доминантты тұқым қуалауға тән белгілер:
Аутосомды-рецессивті тұқым қуалауға тән белгілер:
Х-хромосомамен тіркес доминантты тұқым қуалауға тән белгілер:
Х-хромосомамен тіркес рецессивті тұқым қуалауға тән белгілер:
Голандриялық немесе У-хромосомамен тіркес тұқым қуалауға тән белгілер:
Митохондриялық тұқым қуалауға тән белгілер:
Белгілердің тіркес тұқым қуалауы.
Сызба – 2. Талдаушышағылыстыру №1.
Сызба – 3. Талдаушы шағылыстыру № 2
Нуклеин қышқылдары, құрылысы, жіктелуі
РНҚ –ның өмір сүруге қабілеттілігі:
Эукариоттардың генінің молеклалық құрылымы(схема түрінде)
Ген, классификациясы және функциясы
Геннің атқаратын қызметіне байлансыты гендер екі топқа бөлінеді:
Ағзалардың тіршілік қабілетіне байланысты гендер былай жіктеледі:
Нуклейн қышқылдары мен геннің медициналық маңызы
Сабақ 9 Ақпараттық дидактикалық блок
Репликацияның негізгі ферменттері:
Прокариотгендерініңактивтілігініңреттелуі:
Эукариоттар гендерінің экспрессиясының реттелуі
Гендерактивтілігініңгеномдықдеңгейде(ДНҚ)реттелуі
Эукариоттардың ДНҚ фракцияларының сипаттамалары
Кейбіргендердіңконститутивтік(тұрақты)экспрессиясы
Гендер активтілігінің транскрипциялық деңгейде реттелуі
Супрессор гендердің мутациялары құрылымы мен қызметі бұзылған ақуыздардың синтезделуіне және түрлі мүшелер мен жүйелердегі ісіктік трансформацияның басталуына әкеледі.
Қазіргі уақытта 20-ға жуық ісіктің супрессорлық гендері анықталған. Олардың мутациялары ісіктің дамуына алып келеді. Супрессорлық гендерде жиі кездесетін мутациядардың түрлері - геннің кодтаушы бөлігінде жүретін миссенс мутациялар және нүктелік делециялар.
p53 және Rb антионкогендері жақсы зерттелген.
Ретинобластомада (жиілігі — 20 мың баланың ішінде біреуінде) Rb генінің қызметінің тоқтауы байқалады. 60% жағдайда ретинобластома спорадикалды түрде дамиды, ал 40% жағдайда аутосомды-доминантты түрде тұқым қуалап беріледі. Rb генінің тұқым қуалайтын ақаулығы жағдайында, екінші аллелі қалыпты, сондықтан ісіктің дамуы тек екінші (қалыпты) аллелде қосымша мутация болған жағдайда жүруі мүмкін. Спонтанды түрде дамитын ретинобластомада Rb геннінің қызметінің тоқтауы екі алеллді де бірдей қамтиды (3-сурет).
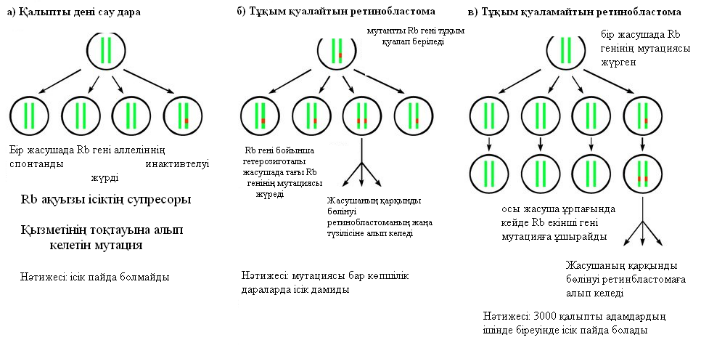
3-сурет. Ретинобластоманың тұқым қуалайтын және тұқым қуаламайтын түрлерінің даму механизмдері.
Қалыпты жағдайда Rb гені ретинбластоманың супрессоры болып, оның дамуын тежейді.
Ісіктің ең маңызды суресорларының бірі р53 (массасы 53 илодальтон, кД) гені болып табылады. Оның көптеген (3400 астам) аллелдері бар. Адамдардағы барлық рактың жартысына жуығы р53 генінің мутантты аллелдеріне байланысты болады.
р53 гені ДНҚ қызметі және құрылымы бұзылған жағдайда, жасушалардың бағдарланған өлімі- апоптозбен аяқтайталатын сатылы механизмдерді іске қосады.
Р53 генінің мутациясы өзгерген жасушаларда апоптоз үрдісінің дұрыс жүрмеуімен олардың тірі қалып, пролиферациялануына, яғни рак жасушасына трансформациялануға алып келеді.
Сүт безі рагінің гендері BRCA1 жәнеBRCA2 де супрессорлық гендер болып табылады. Бұл гендердің мутациялары әйелдерде сүт безі рагінің тұқым қуалайтын түріне және жұмыртқа безінің рагіне алып келеді.
Аталған, онкогенезге алып келетін гендермен қатар, ДНҚ репарациясы гендерін де айта кету керек. Бұл гендер жүйесіндегі өзгерістер жасушаның гендік мутацияларға бейімділігін арттырып, ісіктік үдерістің басталуына негіз болатын, геномның тұрақсыздығына алып келеді. Аутосомды-рецессивті тұқым қуалайтын гендік ауру-пигменттік ксеродерма, ДНҚ репарациясының бұзылыстарының ісік үдерістерімен байланыстылығын көрсетеді.
Қалыпты жасушалық штаммдармен салыстырғанда аномальды рак жасушаларында телофераза ферменті белсенді болғандықтан, тіршілігін жоймайды және қартаймайды. Адамдардағы онкомаркер ретінде аталған ферменттің белсенділігін анықтауға болады. Теломеразаның белсенді болуы, рак жасушаларының пролиферациясына қажет болуы мүмкін.
Канцерогенез себептерін зерттеуде вирустар үлкен орын алады. Адамдарда белгілі бір рак типтерінің дамуы ДНҚ-лы вирустармен байланысты.
Адамдарда рактың дамуына алып келетін ДНК-лы вирустарды 4 топқа жіктеуге болады:
1. Папилломалардың (папилломавирустар) дамуына алып келетін вирустар. Папилома вирусы жатыр мойны рагіне алып келуі мүмкін;
2. Лимфоманың дамуына лаып келетін Эпштайн-Барр (EBV) вирусы;
3. Бауыр рагіне алып келуге қабілетті гепатит В (HBV) вирусы;
4. Капоши (KSHV) саркомасына алып келетін герпес вирусы.
ДНҚ-лы вирустардың ісікке алып келу механизмдері өзгеше болады. Оның геномы, нақтырақ айтсақ, жеке гендері - онкогенді папова вирустардың өнімдері жасушаның бөлінуін тежейтін, пролиферацияның реттелуіне қатысатын ақуыздармен байланысып, алардың белсенілігін тоқтату арқылы жасушаның бақылаусыз бөлінуіне жағдай жасайды. ДНҚ-лы вирустардың трансформациялық белсенділігі олардыңң ие жасушасының геномына ене алу мүмкіндігімен байланысты. Вирустың жасуша геномына енген жері тұрақты болады. Бұл ісіктің шығу тегінің моноклоналды болуын анықтайды.
РНҚ-лы вирустар — ретровирустар. Ретровирустарда автономды репликацияға қажетті гендері болмайды. Сондықтан, вирустар жасушаға енгенен кейін иесінің ақуыздар жүйесін қолданады. Вирустар көбеюді бастау үшін ретровирустардың геномында міндетті түрде болатын ерекше геннің өнімі кері транскриптаза ферментін пайдаланады. Осы ферменттің көмегімен РНҚ-лы вирустар екі тізбекті ДНҚ молекуласына айналап, иесінің геномына ену арқылы өз гендері сияқты қызмет атқара бастайды. Нәтижесінде жаңа вирустардың түзілуіне қажетті ақуыздар мен РНҚ-лар синтезделеді.
Сондай-ақ, кейбір ретровирустарда иесінің жасушасын трансформациялауға қабілетті ерекше вирустық онкогендер деп аталатын гендері болады.
Ең алғаш онкоген - Srс гені 1981 жылы Раус саркома вирусында табылған. Бұл жаңалық жасушаның кейбір вирустармен зақымдалуынан олардың ісікке трансформациялану механизмін түсіндіруге мүмкіндік берді. Ісік жасушаларының дамуы вирустардан емес олардың геномындағы онкогендерден екендігі белгілі болды. Ісіктік вирустар өздігінен ракке алып келе алмайтындығы белгілі болды. Олар жасушаның генетикалық аппаратына онкогенді енгізу арқылы қатерлі ісіктің қалыптасуына алып келеді.
Ісіктер жасушалардың бақыланусыз бөлінуімен сипатталатын, гендік аурулардың ерекше тобы болып табылады. Ағзада таралу қабілетіне қарай оларды екі топқа бөледі: қатерсіз және қатерлі.
Қатерсіз ісік (фиброма, липома, миома, ангиома, невринома, хондрома, остеома, папиллома, аденома, базалиома және т.б.) өсуі баяу және өз ұлпасының аймағынан шықпайды, көрші ұлпаларға енбейді, инвазивті өспейді және метастаз бермейді. Ісік өсінділерінде тамырлар болмайды, сондықтан қорек жеткіліксіз болғандықтан жасушалар тоқтаусыз бөлініп, көбеюіне қарамастан, қартайған жасушалардың өлуі арқылы бұл үрдіс реттеліп отыруы нәтижесінде ісік өспейді.
Қатерлі ісік дегеніміз (фибросаркома, липосаркома, лейомиосаркома, аденокарцинома, тегіс жасушалы рак, хорионкарцинома және т.б.) белгілі ұлпарға еніп өсуге (инвазивті) қабілетті және екіншілік ісіктердің (метастазға) пайда болуына алып келетін, ағзаның басқа бөліктеріне орын ауыстыру қабілетеріне ие.
Рак жасушаларының қасиеттері:
1) Рак жасушаларының негізгі қасиеті оларда жасушаның бөлінуін тежеуші механизмдер мен факторлардың болмауына байланысты тоқсаусыз және бақыланусыз бөлінуі. Кез-келген рак шексіз өсуге қабілетті.
2) Автономдылығы –ісіктер көрші ұлпаларға тәуелсіз болады. Қалыпты даму кезінде ұлпалардың бір-бірімен өзара әсерлесіп, өз аймақтарынан шықпайды. Қатерлі ісіктер бұл әсерді сезбейді.
3) Қатерлі ісіктің келесі қасеиті – жасушалардың тіршілігін жоймауы, ал қалыпты жасушалардың тіршілік циклі бағдарланған өлім- апоптозбен аяқталады. Ісік жасушаларының бөліну шегі ағзада да және ағзадан тыс та жүре береді.
4) Қатерлі ісіктің маңызды және міндетті белгісі моноклоналдығы. Қатерлі ісік генетикалық өзгерген бір жасушадан дамиды. Оның ұрпақтары осы өзгерген бір жасушалардың көшірмесі (клондары) болып табылады.
5) Метастаз беру –бұл өзі метастаздарды қалыптастыру үрдісі. Ісік өсудің нәтижесінде оның жеке жасушалары бөлініп, қанға, лимфаға түсіп, басқа тіндерге тарайды. Онда екіншілік (жаңа) ісіктерді түзеді. Бұл метастаздардың құрылымында әдетте бастапқы пайда болған ісіктен айырмашылық болмайды. Тек қатерлі ісік ғана метастаз береді, ал қатерсіз ісік метастаз бермейді.
Канцерогенез сатылары
Канцерогенез үдерісінде үш негізгі сатыны бөліп көрсетуге болады: инициация, промоция және прогрессия.
1. Инициация. Барлық ісік, жеке жасуша ДНҚ-сының зақымдалуынан басталады. Бұл генетикалық өзгерістер канцерогенді немесе онкогенді вирустар арқылы болуы мүмкін. Бірақ, ісіктің инициациясында протоонкогендердің зақымдалуы маңызды. Бұл зақымдалу сомалық жасушаның трансформацияланып, ісікке айналуында маңызды болып табылады. Ісіктің инициациясына, сонымен қатар антионкогендердің (онкосурпессор гендер) зақымдалулары да алып келуі мүмкін.
2. Ісіктің промоциясы – бұл ісікке алып келетін фактормен зағымдалған жасушаның бөлінуі. Бұл үрдіс жылдар бойы созылуы мүмкін.
3. Ісіктің прогрессиясы — бұл қатерлі ракке алып келетін өзгерген жасушалардың қарқынды бөлінуі, инвазивті өсуі және метастаз беруі.
Қорыта келгенде, рак қарт және егде адамдарда дамиды. Себебі гендерде кездейсоқ пайда болған мутациялар, көп жылдар бойы жинақтап, қажетті деңгейге жеткенде рак ауыруына алып келеді.
Ағзаның иммундық жүйесінің жас өткен сайын әлсіреуі нәтижесінде өзгерген жасушалар тірі қалып, рактың пайда болуына себеп болады.
14 сабақ. Жасушаның бағдарланған өлімінің (апоптоздың) молекулалық-генетикалық механизмдері
Жасушалар белгілі бір уакыт аяғында қызмет атқарады және олардың тіршілігі өліммен аяқталады. Сомалық жасушалардың бөліну саны алдын-ала бағдарланған, шектеулі.
Барлғы болмаса да, көпшілік жасушалар ағзаға қажетсіз болғанда немесе көп зақымдалған жағдайларда ішкі генетикалық бағдарламалардың активтелуі нәтижесінде өзін-өзі жою қабілетіне ие болады. Мұндай жасушаның өлуі бағдарламасының іске асуы морфологиялық өзгерістермен сипатталады. Бұл жасушаның жойылу түрі апоптоз (жапырағынан айырылған өсімдікті сипаттау үшін қолданылатын грек терминінен шыққан) деп аталады.
Жасушаның бағдарланған өлімі барлық тірі ағзаларға, соның ішінде адамға да тән. Күніне адам ағзасында бірнеше ондаған миллиардтаған жасушалар жойылып отырады. Жойылған жасушалардың орны бөліну нәтижесінде (митоз) пайда болған жас жасушалармен толықтырылып отырады.
Бағдарланған жасуша өлімінің екі типін ажыратады: «ішкі» апоптоз және «бұйрық бойынша» апоптоз.
«Ішкі» апоптоз - жасушаның өзіне деген қанағаттанарлықсыз жағдайынан болады.
а) хромосомалардың қатты зақымдалуы (репарацияланбайтын немесе нашар репарацияланатын): хромосомалардың дұрыс сегрегацияланбауы, ДНҚ конформациясын бұзатын көптеген үзілулер, тізбектер арасындағы тігілулер және т.б.
б) жасуша ішілік мембраналардың (митохондриялардың), оның липидтерінің күшті тотықтырғыштар нәтижесінде зақымдалуы.
«Ішкі» апоптоз жасушаның генетикалық материалы, органоидтары зақымдалғанда және қартайғанда іске қосылады. Апоптозға алып келетін жасушаның құрылымының бұзылары өте күшті болмауы керек.
«Бұйрық бойынша» апоптоз мембрана немесе жасуша ішілік рецепторлар арқылы берілетін сыртқы «негативті» синал арқылы іске асады.
Жалпы алғанда жасуша тіршілікке қабілетті, бірақ ағза тұрғысынан қарағанда, ол қажетсіз тіпті қауіпті болуы мүмкін. Апоптоздың бұл түрі онтогенездің белгілі бір кезеңдерімен байланысты: эмбриогенезде бірқатар негіздердің және пронефростың редукциялануы, эмрионалдық дамудың морфогенезінде саусақ аралық «жарғақшалардың» жойылуы.
«Бұйрық бойынша» апоптоз иммундық жүйенің қалыптасуы мен қызмет атқаруында да маңызды: Т және В лимфоциттерінің аутоиммундық клондарының жойылуы.
Әйел адамдардың репродуктивті жүйесінде: атрезиялық фолликулалар жасушаларының (ооцит және фолликулалық жасуша), редукцияланған сары дене жасушаларының, менструация алдында энометрияның функционалдық қабатының, лактация аяқталғаннан кейін сүт безінің лактоциттерінің жойылуы және т.с.с.