ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 04.12.2023
Просмотров: 763
Скачиваний: 1
СОДЕРЖАНИЕ
Сабақ 2 Жасушаның генетикалық аппараты
Сабақ 3 . Тақырып: Сомалық және жыныс жасушалардың бӛлінуі. Тақырыптың негізгі сұрақтары:
Постмитоздық кезең (пресинтездік) кезең G1:
Гаметогенез (оогенез, сперматогенез), сатылар сипаттмамасы. Мейоз анықтамасы және
Митоз және мейоздың медициналық маңызы.
2 сурет. Көз бұршағының индукциясы.
Онтогенездің жасушалық механизмдерi.
Даму ақауларының сызбасы ДТБА жіктелуі:
Сабақ 5 АҚПАРАТТЫҚ – ДИДАКТИКАЛЫҚ БЛОК
Аутосомды-доминантты тұқым қуалауға тән белгілер:
Аутосомды-рецессивті тұқым қуалауға тән белгілер:
Х-хромосомамен тіркес доминантты тұқым қуалауға тән белгілер:
Х-хромосомамен тіркес рецессивті тұқым қуалауға тән белгілер:
Голандриялық немесе У-хромосомамен тіркес тұқым қуалауға тән белгілер:
Митохондриялық тұқым қуалауға тән белгілер:
Белгілердің тіркес тұқым қуалауы.
Сызба – 2. Талдаушышағылыстыру №1.
Сызба – 3. Талдаушы шағылыстыру № 2
Нуклеин қышқылдары, құрылысы, жіктелуі
РНҚ –ның өмір сүруге қабілеттілігі:
Эукариоттардың генінің молеклалық құрылымы(схема түрінде)
Ген, классификациясы және функциясы
Геннің атқаратын қызметіне байлансыты гендер екі топқа бөлінеді:
Ағзалардың тіршілік қабілетіне байланысты гендер былай жіктеледі:
Нуклейн қышқылдары мен геннің медициналық маңызы
Сабақ 9 Ақпараттық дидактикалық блок
Репликацияның негізгі ферменттері:
Прокариотгендерініңактивтілігініңреттелуі:
Эукариоттар гендерінің экспрессиясының реттелуі
Гендерактивтілігініңгеномдықдеңгейде(ДНҚ)реттелуі
Эукариоттардың ДНҚ фракцияларының сипаттамалары
Кейбіргендердіңконститутивтік(тұрақты)экспрессиясы
Гендер активтілігінің транскрипциялық деңгейде реттелуі
Прокариотгендерініңактивтілігініңреттелуі:
Транскрипциялық бақылаудың екі түрі бар: негативті және позитивті.
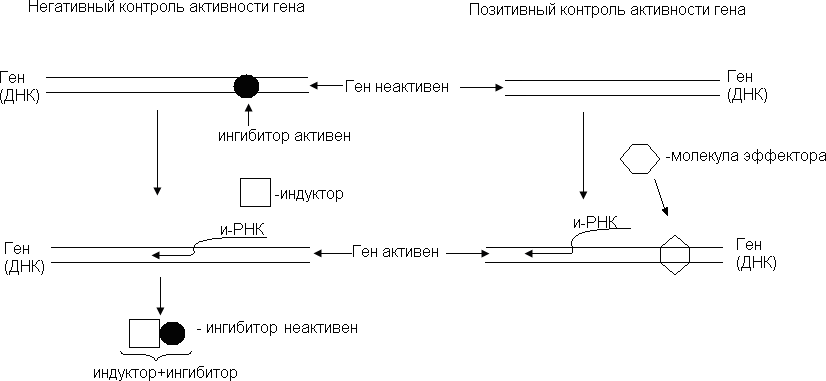
Ген активтілігін негативті бақылау жасушада геннің транскрипциясын тоқтататын арнайы факторлар – ингибиторлардың болуына негізделген. Жасушада сонымен қатар
транскрипцияның басталуына мүмкіндік беріп, ингибиторларға қарсы әрекет көрсететін индуктор-факторлары болады.
Ген активтілігін позитивті бақылау, промоторды активтендіріп, транскрипцияны бастайтын эффекторлық ақуыз молекулалары мен геннің әрекеттесуіне байланысты.
Көпшілік жағдайда бақылаудың осы екі түрі де қатарынан жүреді. Мысалы, катаболистік (ыдыратушы) жүйелер позитивтік жолмен де, негативтік жолмен де реттелуі мүмкін.
Негативтікбақылау
Индукцияланатынжүйелер(индукцияланатыноперондар).Бұл жүйелерде ферменттіңсинтезі үшін жасушаны қоршаған ортада индуктор деп аталатын арнайы субстрат-затының болуы қажет.
Алғаш рет Ф. Жакоб және Ж. Моно (1961) индуцибельді жүйені тәжірибе жүзінде зерттеу негізінде ішек таяқшасындағы (Е.coli)β – галактозидаза ферментінің синтезінің реттелуін Lac – оперон моделін ұсыну арқылы сол ферменттердің синтезделуінің индуцибельді (қоздырушы) немесе репрессибельді (басылыңқы) механизмдерін түсіндірді.
Жакоб және Моно бойынша Lac – оперонының құрылысы мен қызметі
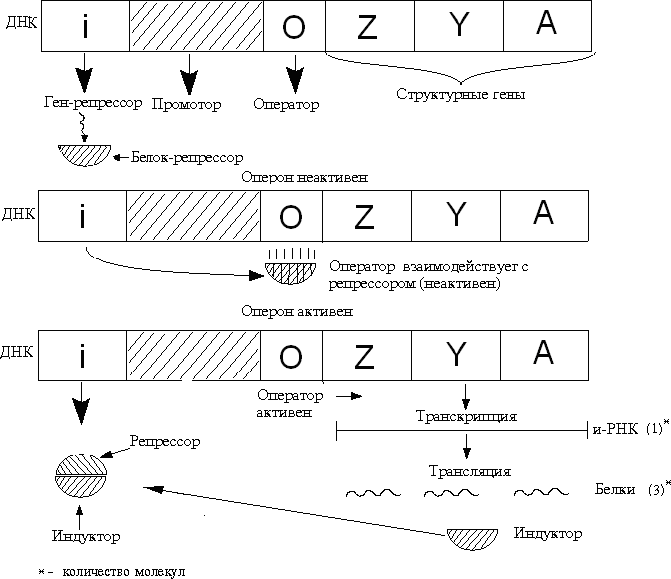
Лактозалық оперонныі сызбасы
Оперон
ақуызсинтезініңбақылануыныңструктуралық-функционалдықбірлігініңэлементтері:
-
Құрылымдық гендер - ферменттер синтезіне қатысады -
Оперон - белок-репрессоры байланысатын сайт
3.Промотор - РНК-полимераза байланысатын сайт
Оперон құрамындағы қатарласа орналасқан құрылымдық гендер - кластерлық гендер деп аталады. Осы кластерлық гендерден тұтас бір ғана полицистронды РНҚ түзіледі. Полицистронды РНҚ-дан трансляция нәтижесінде жалпы қызмет атқаратын бірнеше белок-ферменттер синтезделеді.
Лактозалық оперондағы (Lac – оперон) құрылымдық гендер үш цистроннан тұрады: Z,Y,A. Бұл гендердің түзілетін өнімі – (ферменттер) лактоза қантын ыдыратуға қатынасады, β-галактозидаза ферментін синтездейтін Z – гені лактозаны глюкоза мен галактозаға дейін ыдырады. Y генінің өнімі – фермент галактозид-пермеаза. Бұл ферменттің қызметі жасушаның мембранасымен байланысып лактозаның жасушаға сіңірілуін қамтамасыз етеді. А-генің өнімі – тигалактозид – ацетилаза ферменті лактозаның толық ыдырауына қатынасады.
Қалыпты жағдайда, яғни жасушада лактоза жоқ кезде бұл үш ферменттердің синтезі жүрмейді, осы аталған ферменттердің аздаған молекулалары ғана болуы мүмкін.
Жасушаға лактоза сіңірілген кезде барлық үш ферменттің синтезі қарқынды жүре бастайды. Олай болса, лактозаның жасушаға енуі оны ыдыратып пайдалануға қажетті ферменттердің синтезін индукциялайды. Барлық үш фермент бір ғана полицистрондық а- РНҚ-ның трансляциясына байланысты түзілгендіктен бұл жағдайды жасушаға лактоза – индуктордың енуіне жауап ретінде оперонның біртұтас үйлесімді
қызметі деп түсіндіреді.
Жакоб және Моно теориясы бойынша реттеуші ген (i) репрессор-ақуызын синтездейді, ортада индуктор (лактоза) жоқ кезде ол оператормен (о) байланысып оны активсіздендіреді, Lac –оперондағы үш Lac –цистрондар транскрипцияланбайды. Құрылымдық гендердің транскрипциясының индукциясын (басталуы) былай түсіндіруге болады, жасушаға енген лактоза индуктор ретінде репрессор белогымен байланысып оны оператордан ажыратып активсіздендіреді. Репрессордан ажыраған оператор оперонның жұмыс істеуіне мүмкіндік береді: құрылымдық гендердің үйлесімді транскрипциясы мен трансляциясы басталады.
Репрессор белогында екі түрлі қайта жабылмайтын сайттар болады: 1) оператормен байланыстыратын сайт; 2) индуктормен байланысу сайты. Жакоб және Мононың зерттеулері кезінде (1961 жылдары) промотордың бар екендігі және репрессорлық реттелудің механизмі жайлы деректер болмаған. Кейінірек E. coli-дің ДНҚ тізбегінде РНҚ полимераза ферменті жалғанатын промоторлық сайттың бар екндігі және ол сайт репрессор (і) гені мен оператор (о) гендерінің арасында орналасқаны белгілі болды.
Репрессор гені Lac – оперонынан бөлек моноцистронды а-РНҚ транскрипциаланып репрессор ақуызын синтездейді. Ағзаға түскен индукторға жауап ретінде барлық үш фермент бір ғана полицистронды а-РНҚ трансляциялануынан синтезделетін болғандықтан оперон бір тұтас жүйе ретінде жұмыс істейді.
Позитивтікбақылау
Көптеген бактериялық гендер позитивтік бақылаудың қарамағында болады. Гендер активтілігінің позитивтік бақылану механизмі нақты эффектор факторы бар кезде активтілікті күшейтіп РНҚ-полимеразаның промоторға жалғануы мен а-РНҚ-ның синтезделуін (транскрипцияны) жылдамдатады.
Гендер активтілігін позитивті бақылауға мысал ретінде өзімізге белгілі E.coli-дің Lac
–оперонын алайық. Позитивтік бақылаудың мәні мынада, жасушада глюкоза мен галактоза бар кезде Lac-оперон активті емес, сондықтан Lac – а-РНҚ-ның синтезі жүрмейді, β- галактозидаза ферменті де синтезделмейді. зделмейді
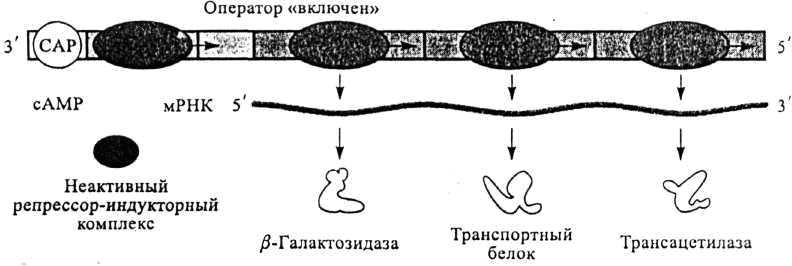
lac-оперонның позитивті реттелуі
Лактозалық і-репрессорын активсіздендіру үшін ортада тек глюкозаның болғаны жеткіліксіз. Глюкозаға қосымша элемент циклдық АМФ-катаболизмдік активтендіруші белок (КАБ)-тың болуы қажет. Олай болса жасушада Lac – а-РНҚ-ның транскрипциясы ол ортада индуктор ретінде глюкоза және циклдық (ц)АМФ-КАБ бар кезде жүреді. Циклдық (ц) АМФ-КАБ –тың активтілігі глюкозаның концентрациясына байланысты. Глюкоза Lac
– оперонның активтілігіне жанама ингибиторлық (тоқтататындай) әсер етеді.
Циклдық АМФ-тың кішігірім молекулалары ішек таяқшасынан басқа көптеген бактерияларда да кездеседі, олардың синтезделуі аденил – циклаза ферментінің көмегімен жүреді, оның жасушадағы концентрациясы жанама түрде глюкозаның метаболизмімен реттеледі. Егерде бактерия өсетін ортада глюкоза жеткілікті болатын болса онда цАМФ концентрациясы төмен болады, ал ортада глюкозаның метаболизміне қатыспайтын
глицерол немесе көмірсулардың басқада көзідері болатын болса цАМФ концентрациясы жоғарылайды. цАМФ концентрациясының бақылану механизмі толық ашылған жоқ.
Lac – оперонының активтілігінің реттелуінде цАМФ маңызды. Сонымен қатар ішек таяқшасының жасушасында Lac – оперонының активтілігін реттеуге қатынасатын катаболизмдік активтендіруші белок (КАБ) «сrp» генінің өнімі болып табылады. Осы сrp немесе аденил – циклаза гендерінің мутацияға ұшырауы Lac – а-РНҚ-ны синтездеу қабілетінен айырады, сондықтан Lac а-РНҚ синтезінің басталуы үшін жасушада осы екі геннің (цАМФ және КАБ) өнімдерінің болуы қажет. КАБ және ц АМФ бірігіп бірге Lac- жүйесінің активті реттеуші элементі болып табылады. Олар транскрипцияның басталуы үшін ДНК тізбегінің промоторлар қатарымен байланысуы қажет.Олай болса цАМФ және КАБ кешені репрессорға (негативті реттелуге) қарама-қарсы қасиет көрсететін позитивтік реттеуші болып табылады. Бұл Lac- оперонының активтілігі бір-біріне тәуелсіз позитивті және негативті механизмдермен реттелетіндігін көрсетеді.
Репрессибельдікжүйелер
Жасушадағы кейбір ферменттердің мөлшері олар синтездейтін соңғы өнімдердің концентрациясына байланысты өзгеріп отырады. Егер жасушада соңғы өнімдердің мөлшері жоғарылап, артып кетсе ол ферменттердің синтезі тоқталады. Ондай ферменттерді репрессибельдік ферменттер деп, ал соңғы өнімдерін – корепрессор деп атайды.
Реттеуші ген апорепрессор деп аталатын затты синтездейді. Апорепрессор корепрессормен қосылып активті қызмет атқаратын репрессор молекуласын түзейді. Репрессор синтездік үдерістерге қатынасатын барлық гендердің активтілігін тежейді.
Мысалы, Salmonella typhimurium бактериясында гистидин аминқышқылының концентрациясы жоғары ортада гистидин корепрессор ретінде әсер етеді. Ол корепрессор ретінде