ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 04.12.2023
Просмотров: 759
Скачиваний: 1
СОДЕРЖАНИЕ
Сабақ 2 Жасушаның генетикалық аппараты
Сабақ 3 . Тақырып: Сомалық және жыныс жасушалардың бӛлінуі. Тақырыптың негізгі сұрақтары:
Постмитоздық кезең (пресинтездік) кезең G1:
Гаметогенез (оогенез, сперматогенез), сатылар сипаттмамасы. Мейоз анықтамасы және
Митоз және мейоздың медициналық маңызы.
2 сурет. Көз бұршағының индукциясы.
Онтогенездің жасушалық механизмдерi.
Даму ақауларының сызбасы ДТБА жіктелуі:
Сабақ 5 АҚПАРАТТЫҚ – ДИДАКТИКАЛЫҚ БЛОК
Аутосомды-доминантты тұқым қуалауға тән белгілер:
Аутосомды-рецессивті тұқым қуалауға тән белгілер:
Х-хромосомамен тіркес доминантты тұқым қуалауға тән белгілер:
Х-хромосомамен тіркес рецессивті тұқым қуалауға тән белгілер:
Голандриялық немесе У-хромосомамен тіркес тұқым қуалауға тән белгілер:
Митохондриялық тұқым қуалауға тән белгілер:
Белгілердің тіркес тұқым қуалауы.
Сызба – 2. Талдаушышағылыстыру №1.
Сызба – 3. Талдаушы шағылыстыру № 2
Нуклеин қышқылдары, құрылысы, жіктелуі
РНҚ –ның өмір сүруге қабілеттілігі:
Эукариоттардың генінің молеклалық құрылымы(схема түрінде)
Ген, классификациясы және функциясы
Геннің атқаратын қызметіне байлансыты гендер екі топқа бөлінеді:
Ағзалардың тіршілік қабілетіне байланысты гендер былай жіктеледі:
Нуклейн қышқылдары мен геннің медициналық маңызы
Сабақ 9 Ақпараттық дидактикалық блок
Репликацияның негізгі ферменттері:
Прокариотгендерініңактивтілігініңреттелуі:
Эукариоттар гендерінің экспрессиясының реттелуі
Гендерактивтілігініңгеномдықдеңгейде(ДНҚ)реттелуі
Эукариоттардың ДНҚ фракцияларының сипаттамалары
Кейбіргендердіңконститутивтік(тұрақты)экспрессиясы
Гендер активтілігінің транскрипциялық деңгейде реттелуі
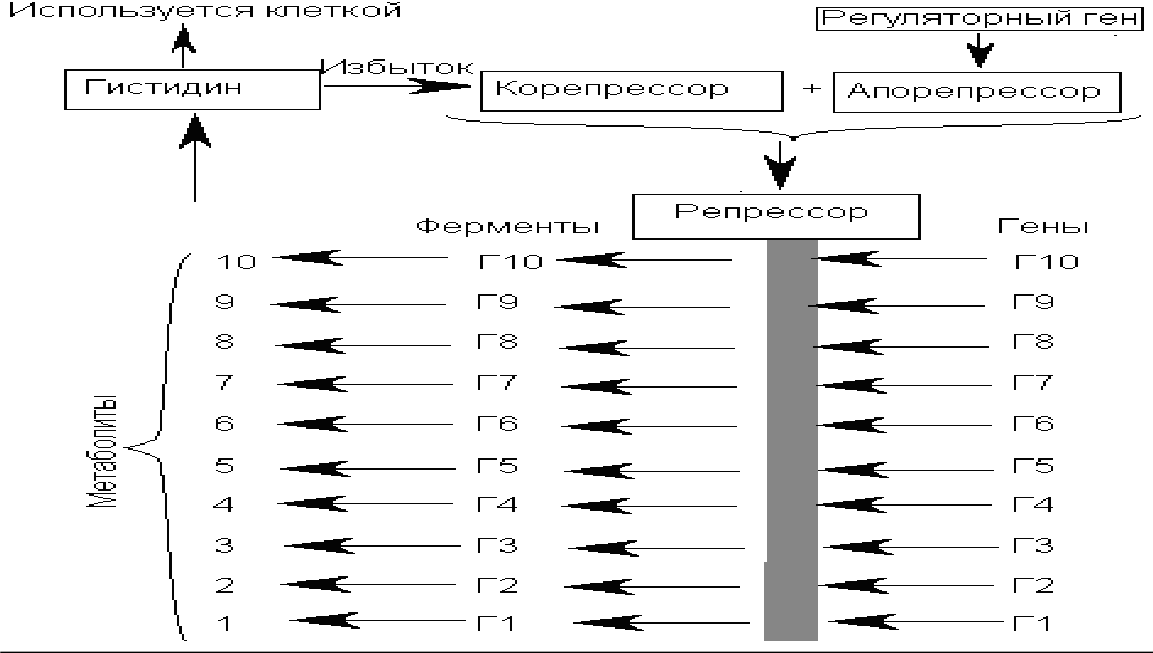
Мұндай репрессия типін үйлесімдік (координациялық) репрессия деп атайды.
Репрессияның бұл типін координациялау репрессиясы деп атайды. Salmonellatyphimurium-ныңкоординацияланғанрепрессиясыдепатайды.
Трансляциялықбақылау
Прокариоттық гендер активтілігінің трансляциялық деңгейде реттелуі а-РНҚ-ның өмір сүру ұзақтығына тікелей байланысты. А-РНҚ-ның ферменттік ыдырауы оның 51- ұшынан басталып 31-ұшында аяқталады, яғни а-РНҚ-ның бірінші болып синтезделген ұшы, бірінші болып ыдырайды. Ішек таяқшасының а-РНҚ-сының орташа өмір сүру ұзақтығы -2 минутке жуық. А-РНҚ-ның ферменттік ыдырау жылдамдығына оның 51- ұшында орналасқан арнайы нуклеотидтар жүйесі де әсер етуі мүмкін.
А-РНҚ-ның өмір сүру ұзақтығы сонымен қатар оның трансляциясына қатынасатын бос рибосомалардың санына байланысты ауытқиды. Осы себепті бактерияларда түрлі белоктар синтезі жылдамдығының түрліше болуы трансляция жылдамдығымен емес сол жасушадағы рибосомалар санымен анықталады.
Мысалы, ішек таяқшасының (E. coli) лактозалық жүйесіндегі жалпы бір оператормен бақыланатын құрылымдық гендердегі үш цистрон үш ферментті синтездейтіні айтылды. Олар: а) β-галактозидаза; ә) галактозид-пермеаза; б) галактозид-ацетилаза. Бұл белоктардың синтезделу арақатынасы 1:1/2:1/5-ке тең. Бұл арақатынас осы үш белоктың трансляциясы жүретін полицистрондық а-РНҚ-ның 51-ұшынан олардың орналасу арақашықтығын бейнелейді. Гендер неғұрлым полицистронды а-РНҚ - 51-ұшынан неғұрлым алыс орналасқан сайын синтезделетін белоктың мөлшері де азая түседі. Рибосомалардың а-РНҚ-ғы түрлі старттық нүктелерге жалғануы оперондағы құрылымдық Z, Y, A гендері синтездейтін белоктардың сан мөлшерінің түрліше болуына әсерін тигізеді деген жорамал айтылады.
Посттрансляциялықбақылау
Гендер активтілігі белоктардың синтезі аяқталғаннан кейін де реттелуі мүмкін.
Бақылаудың бұл түрін гендер
активтілігінің посттрансляциялық бақылануы деп атайды.
Гендер активтілігінің кері байланысу ұстанымы бойынша басылуы ферменттің синтезіне емес, оның активтілігіне әсер ететін реттеуші механизм болып табылады. Мұнда биосинтез нәтижесінде түзілген концентрациясы жоғары соңғы өнім бірінші синтезделген ферментпен байланысып метаболизмдік үдерістің жүруіне жағдай туғызады.
Соңғы өнім + бірінші фермент кешені ферменттің төртінші реттік құрылымын өзгертіп, оның катализдік сайтын активсіздендіреді. Фермент құрылымының өзгеруі оның
өзінің активтілігін тоқтатады және соңғы өнім мен аралық ыдырау заттарының артық мөлшерде синтезделуіне кедергі жасайды. Мысалы, ішек таяқшасында изолейциннің синтезделу үдерісін зерттеу барысында (Umbеrger, 1961) изолейцинді (треонин алмасудағы соңғы өнім) жасанды ортаға енгізгенде, сол мезетте треониннің изолейцинге айналу үдерісі тоқталған. Изолейциннің мөлшері шамадан тыс артқан жағдайда жасушада бұл аминқышқылының синтезі тоқталып, экзогендік изолейцинді пайдалану басымырақ жүрген.
Треониннің изолейцинге айналу процесіне қатынасатын 5 фермент артық мөлшердегі изолейцинмен өзара әрекеттесуі барысында олардың әрқайсысының синтезі де тоқталады.
Эукариоттар гендерінің экспрессиясының реттелуі
Прокариоттарға қарағанда эукариоттардың құрылымы күрделі. Мысалы, адам ағзасында 200 астам әртүрлі жасушалар және 100 ақуыздар болады. Эукариоттар гендерінің экспрессиясының реттелуінің тек өзіне тән бақылану механизмдері бар.
Эукариоттарда гендер активтілігі реттелуінің екі типін ажыратады:
-
қысқа мерзімді немесе қайтымды реттелу бактериялардағы реттелуге сәйкес, ортадағы өзгерістерге жасушаның жауап қайтаруы болып табылады. Реттелудің бұл типі жасушадағы субстрат деңгейінің жоғары не төмен болуына байланысты ферменттер активтілігнің өзгеруімен сипатталады. Осы реттелу типіне сонымен қатар, митоздың түрлі кезеңдерінде байқалатын ДНҚ, РНҚ және белоктардың синтезделу жылдамдығының өзгерістерін де жатқызады. -
Ұзаққа созылған немесе қайтымсыз реттелу – дамудың ұрықтану және зигота түзілу кезеңінен бастап көп жасушалы ағза түзілуге дейінгі аралықта жүретін детерминация, дифференциация т.б. үдерістерінде жүреді.
Эукариоттарда гендер активтілігі реттелудің осы екі типі тұқым қуалау ақпаратының жүзеге асырылуының мына кезеңдерінде байқалады:
-
Геномдық деңгейде (ДНҚ) реттелу; -
Транскрипциялық деңгейде реттелу; -
Посттранскрипциялық деңгейде реттелу -
Трансляциялық деңгейде реттелу; -
Посттрансляциялық деңгейде реттелу;
Гендерактивтілігініңгеномдықдеңгейде(ДНҚ)реттелуі
Эукариоттар геномында нуклеотидтер көп, тек оның 2-5% ДНҚ-сы ғана ақуызды кодттайды. Адам геномында 3,3 млрд. жуық нуклеотидтер жұбы (гаплоидты жиынтықта) болады, соның ішінде 3% кодтаушы аймақтың үлесіне тиеді.
Эукариоттар геномының ерекшелігі де сол,
ДНҚ кодтаушы және реттеуші (сигналдық) қатарлары бола тұра көптеген сайттары транскрипцияланбайды. Барлық сома жасушаларының геномдары бірдей бола тұра әртүрлі типті жасушаларда әртүрлі гендер экспессияланады. Бұндай механизмнің болуы тіршілік барысында тұрақты түрде бір гендердің экспрессиялануын және екінші бір гендердің экспрессиясының тежелуін қамтамасыз етіп отырады.
Эукариоттардың ДНҚ фракцияларының сипаттамалары
| ДНҚфракциятүрлері | жалпы ДНҚ санына шаққандағ ы ДНҚ %мөлшері | Геномдағы кездесу жиілігі | Функционалдық маңызы | Структуралық құрылымы |
| 1. Уникальдысирек қайталанатын қатары | 56-75% | 1 рет | Структуралық және функционалдық гендерден тұрады | Эухроматин |
| 2. Орташа жиілікпен қайталанатын нуклеотидтер қатары | 8-15% | 100-10 000 көшірмелері | Гистонды ақуыздарда кодтаушы гендер, ақуыздар, тРНҚ және рРНҚ | Эухроматин |
| Көпқайталанатын нуклеотидтер қатары | 10-12% | от 10 000 до 1 000 000 көшірмелері |
бөліктері (құрылысытық қызметі)
спейсерлер (гендер арасын бөлуші), экспрессиясын реттейтін гендер (промоторлар, терминаторлар және т.б.) | Гетерохроматин Эухроматин |