ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 04.12.2023
Просмотров: 758
Скачиваний: 1
СОДЕРЖАНИЕ
Сабақ 2 Жасушаның генетикалық аппараты
Сабақ 3 . Тақырып: Сомалық және жыныс жасушалардың бӛлінуі. Тақырыптың негізгі сұрақтары:
Постмитоздық кезең (пресинтездік) кезең G1:
Гаметогенез (оогенез, сперматогенез), сатылар сипаттмамасы. Мейоз анықтамасы және
Митоз және мейоздың медициналық маңызы.
2 сурет. Көз бұршағының индукциясы.
Онтогенездің жасушалық механизмдерi.
Даму ақауларының сызбасы ДТБА жіктелуі:
Сабақ 5 АҚПАРАТТЫҚ – ДИДАКТИКАЛЫҚ БЛОК
Аутосомды-доминантты тұқым қуалауға тән белгілер:
Аутосомды-рецессивті тұқым қуалауға тән белгілер:
Х-хромосомамен тіркес доминантты тұқым қуалауға тән белгілер:
Х-хромосомамен тіркес рецессивті тұқым қуалауға тән белгілер:
Голандриялық немесе У-хромосомамен тіркес тұқым қуалауға тән белгілер:
Митохондриялық тұқым қуалауға тән белгілер:
Белгілердің тіркес тұқым қуалауы.
Сызба – 2. Талдаушышағылыстыру №1.
Сызба – 3. Талдаушы шағылыстыру № 2
Нуклеин қышқылдары, құрылысы, жіктелуі
РНҚ –ның өмір сүруге қабілеттілігі:
Эукариоттардың генінің молеклалық құрылымы(схема түрінде)
Ген, классификациясы және функциясы
Геннің атқаратын қызметіне байлансыты гендер екі топқа бөлінеді:
Ағзалардың тіршілік қабілетіне байланысты гендер былай жіктеледі:
Нуклейн қышқылдары мен геннің медициналық маңызы
Сабақ 9 Ақпараттық дидактикалық блок
Репликацияның негізгі ферменттері:
Прокариотгендерініңактивтілігініңреттелуі:
Эукариоттар гендерінің экспрессиясының реттелуі
Гендерактивтілігініңгеномдықдеңгейде(ДНҚ)реттелуі
Эукариоттардың ДНҚ фракцияларының сипаттамалары
Кейбіргендердіңконститутивтік(тұрақты)экспрессиясы
Гендер активтілігінің транскрипциялық деңгейде реттелуі
Репликация – ДНҚ өздігінен еселенуі, жасушаның бөлінуінің алдында, интерфазаның S кезеңінде өтеді және жасушадағы хромосомалар санының тұрақтылығын қамтамасыз етеді. Хромосомалық ДНҚ мөлшерінің өте үлкен болуына байланысты ДНҚ репликациясы бірден бірнеше жерден басталады (полирепликонды) және екі бағытта көршіліс репликациялық айырдың бірігуіне дейін жүреді. Прокариот ДНҚ-да және
органоидтарында (митохондриялар және пластидтер) бір ғана инициациялық нүкте, бір репликон (монорепликонды) болады. Репликон-ДНҚ репликациясының бірлігі.
Репликация үдерісі мына ұстанымдардың негізінде іске асырылады:
-
жартылай консервативті – ДНҚ-ның бастапқы тізбектерінің әрқайсысы жаңа тізбекке матрица (қалып) болады -
комплементарлық – ДНҚ молекуласының жаңа тізбегі комплементарлық ұстанымға негізделеді, аденин тиминмен, гуанин цитозинмен және керісінше байланысу арқылы түзіледі -
антипараллельдік – ДНҚ-ның жаңа тізбектері бастапқы матрицалық тізбектерге қарама-қарсы бағытта синтезделеді -
униполярлығы - комплементарлы жаңа тізбектердің синтезделуі тек 5¢-3¢бағытында іске асырылады -
үзілмелігі – репликация ДНҚ молекуласының бірнеше жерінде бір уақытта басталады да артынан бір тізбекке тізбектеледі.
Репликация сатылары: инициация, элонгация, терминация.
Репликация инициациясы – ерекше нуклеотидтер қатары бар, инициация нүктесінен, «репликациялық көзшеден» басталады. Сол жерден бастап хеликаза ферментінің көмегімен қосарланған ДНҚ тізбектері тарқатылады, топоизомераза репликацияланбаған ДНҚ бөлігінің алдындығы суперспиральды ажыратады, ал SSB ақуыздары – ажыраған бір тізбекті ДНҚ бөліктерін
тұрақтандырады. ДНҚ синтезі тек бір 5’®3’ бағытында жүреді, ал ДНҚ тізбектері антипараллельді, сондықтанда ДНҚ бір тізбегі үздіксіз синтезделеді және оны лидерлік (бастаушы), екінші тізбекті ілесуші (артта қалушы) тізбек, қысқа кесінділер (Оказаки фрагменттері), түрінде синтезделеді, соңынан бұл кесінділер тізбектеліп жалғанады.
Элогация - ДНҚ репликациясының ұзару үдерісі, ДНҚ жаңа тізбегінің синтезіне бастама беретін РНҚ праймазаның қысқа фрагментті РНҚ праймерлерінің синтезделуінен басталады. Содан кейін РНҚ праймерлерге ДНК-полимераза байланысады да үздіксіз лидерлік тізбек синтезделеді. Ілесуші тізбектің лидерлік тізбектен синтезделу ерекшелігі ДНҚ қысқа, Оказаки фрагменттері түрінде синтезделеді. Ілесуші тізбектің синтезіне бірнеше праймерлер қажет, кейін олар эндонуклеаза көмегімен алынып тасталынады, орны бос қалады. Бос қалған орындар ДНК-полимеразамен көмегімен комплементарлы нуклеотидтермен толықтырылады да тізбектеліп лигаза ферментімен тігіледі.
Терминация - репликация ДНҚ бөлігінің терминациялық сайт деп аталатын арнайы нуклеотидтер қатармен аяқталады. Терминация tus генімен бақыланады, tus генінің өнімі болып табылатын tus ақуызы терминациялық сайтты танып байланысып үдерісті тоқтатады.
Репликация –көптеген ферменттердің қатысуымен жүретін күрделі үдеріс.
Репликацияның негізгі ферменттері:
-
ДНҚ-полимераза – ДНҚ тізбегін синтездейді -
топоизомераза – репликациялық айыр алдындағы ДНҚ-ың суперспирализациясын босаңсытады, тарқатады -
хеликаза -ДНҚ тізбектерін ажыратады -
SSB ақуыздары – ажыраған бір тізбекті ДНҚ бөліктерін тұрақтандырады -
лигаза – ДНҚ фрагменттерін тізбектеп тігеді -
РНҚ-праймаза - ДНҚ- полимераза үшін РНҚ-бастаушты (праймерлер) синтездейді
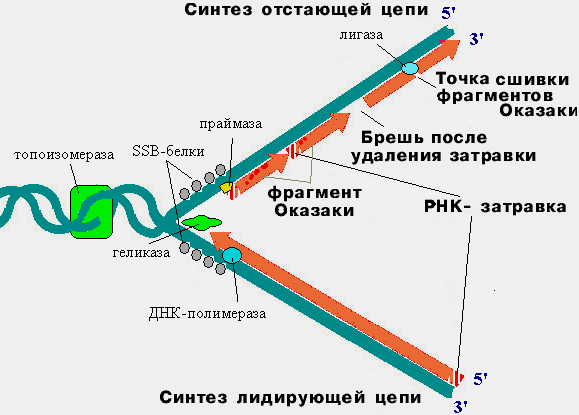
Репликациялық айыр аймағындағы ДНҚ репликацииясының сызбасы
Репликацияның өте жоғары дәлділігі қосымша ДНК-полимеразамен қамтамасыз етіледі. Бұл ферменттің қызметі ДНҚ комплементарлық ұстанымдарын және дұрыс емес нуклеотидтерді алып тастайды түзету және жөндеу жүргізеді.
Прокариоттарда секундына 3000 жұпқа жуық нуклеотидтер, ал эукариоттарда - секундына 100-300 жұп нуклеотидтер синтезделеді.
Дегенмен де репликация кезіндегі гендік (нүктелік) мутациялар – азотты негіздердің алмасуы, репликация қателіктеріне әкеледі. Уотсон және Крик қос спиральды ДНҚ моделінің негізінде ДНҚ-ң азоттық негіздерінің алмасуының алғашқы механизмін ұсынған. Олардың көз қарастары бойынша нуклеотидтер арасындаға комплементарлы A-T және G-C жұптары арасында таутомеризаця жүруі де мүмкін, бұл жағдайда комплементарлық конформациясы өзгереді, аденин гуаниннің қызметіне, гуанин-аденинге, цитозин-тиминге, тимин-цитозинге ауысады. Сонымен қатар репликацияның бұзылу себептеріде әртүрлі болуы мүмкін. Мысалы, ультракүлгін сәуленің әсерінен нуклеотидтер концентрациясының өте
жоғары немесе өте төмен болуы, пуриндік негіздердің спонтанды түсіп қалуы, цитозиннің дезаминденуі себебінен урацилге айнала алады. Пайда болған қателіктер мен бұзылыстар репарациялық үдеріспен және ДНҚ-полимеразаның жөндеу және түзету жүргізе алу қабілетіне байланысты қателіктер түзетіліп отырады, ал кейбір жағдайларда ДНҚ молекуласындаға қателіктер түзетілмей қалып қояды. Егер зақымданулар ДНҚ-ң функциональды активті емес аймағында өтетін болса, онда ондай қателіктер фенотиптік көрініс бермейді және ешқандай зілді қауіптер байқалмайды, бірақ қандайда бір генде нуклеотидтер қатарының жұптасуында қателіктер кететін болса, онда фенотиптік өзгерістің көрініс беру мүмкіндігі көбее түседі де гендік аурулардың пайда болуына әкеледі.
Репликация үдерісіне әртүрлі сыртқы орта факторлары, соның ішінде дәрілік заттарда әсер етеді. Мысалы, дауномицин және ісікке қарсы препараттар ДНҚ құрылысын өзгертеді, нәтижесінде репликация ферменттері жұмыстарын тоқтатады. Мысалы, топоизомераза ДНҚ босаңсыту (деспиралдандыру) мүмкіндігінен айырылады. Алкилдеуші заттар тиофосфамидтер де ДНҚ молекуласындағы азотты негіздердің модификациясын тудырады да репликация ферменттерінің жұмысын тоқтатады. ДНК- топоизомеразасының ингибиторы-новобиоцин ферменттер жұмысына араласып ДНҚ деспиральдануын тоқтатады.
Сонымен репликация үдерісі генетикалық ақпараттың берілуінің негізгі кезеңі, ондағы бұзылыстар жасушаның ары қарай бөлінуін тоқтатуы мүмкін. Сол сияқты репликация жылдамдығының (төмендеуі) өзгеруі мүшелердің дамымай қалуына, эмбриогенезде
репликацияның жоғарлауы мүшенің гиперплазиясына және гендік аурулардың пайда болуына себепші
болады.
ДНҚ молекуласындаға генетикалық ақпарат рибонуклеин қышқылының (РНК) молекуласына беріледі. РНҚ-ң ДНҚ-дан айырмашылығы қант молекуласы дезоксирибозаның орнында рибоза және азотты негізі - урацил (тиминнің орнына) аденинге комплментарлы. ДНҚ транскрипциясы инициация, элонгация және терминации сатыларынан тұратын ДНҚ екі тізбегінің бірінен РНҚ молекуласы синтезделетін матрицалық үдеріс.
Инициация - алғашқы нуклеотид аралық байланыс РНҚ-полимеразаның промоторымен байланысуынан басталады. Осы кезде белгібір жерден ДНҚ тізбегі тарқатылып транскрипциялық «көзше» пайда болады. Прокариоттарда РНҚ-полимераза Прибнов бокстың нуклеотидтер қатарындағы s(сигма) – факторды таниды, соңынан оған екі a тізбегінен, b(бета) b¢(бета штрих) және w (омега) суббірлігінен тұратын кор-фермент қосылады. Бұл кешен холофермент деп аталып транскрипцияның басталу нүктесіне қарай жылжиды, бірнеше нуклеотидтер транскрипцияланғанан кейін, s фактор ферментімен байланысын үзеді, тек қана кор-фермент ДНҚ бойымен жылжиды.
Эукариоттардың РНҚ-полимеразасы өздігінен транскрипцияны инициациялай алмайды, оған жалпы транскрипциялық фактор-ақуызы (ЖТФ) қажет. Ол РНК-полимеразаны ТАТА-бокспен байланыстырады және бұл кешен (РНҚП+ЖТФ) транскрипцияның басталу нүктесіне қарай жылжиды.
Элонгация - РНҚ тізбегінің ұзаруы, транскрипцияның басталу нүктесінен басталып және терминациялық сайтта униполярлы, консервативті және комплементарлы ұстаныммен аяқталатын үдеріс.
Терминация- транскрипцияның аяқталуы, геннің терминациялық сайт қатарындағы ГЦ- ға бай бөлігінде бір тізбекті РНҚ молекуласының бөлініп шығуымен аяқталады. ДНҚ әрбір бөлігінде