ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 04.12.2023
Просмотров: 747
Скачиваний: 1
СОДЕРЖАНИЕ
Сабақ 2 Жасушаның генетикалық аппараты
Сабақ 3 . Тақырып: Сомалық және жыныс жасушалардың бӛлінуі. Тақырыптың негізгі сұрақтары:
Постмитоздық кезең (пресинтездік) кезең G1:
Гаметогенез (оогенез, сперматогенез), сатылар сипаттмамасы. Мейоз анықтамасы және
Митоз және мейоздың медициналық маңызы.
2 сурет. Көз бұршағының индукциясы.
Онтогенездің жасушалық механизмдерi.
Даму ақауларының сызбасы ДТБА жіктелуі:
Сабақ 5 АҚПАРАТТЫҚ – ДИДАКТИКАЛЫҚ БЛОК
Аутосомды-доминантты тұқым қуалауға тән белгілер:
Аутосомды-рецессивті тұқым қуалауға тән белгілер:
Х-хромосомамен тіркес доминантты тұқым қуалауға тән белгілер:
Х-хромосомамен тіркес рецессивті тұқым қуалауға тән белгілер:
Голандриялық немесе У-хромосомамен тіркес тұқым қуалауға тән белгілер:
Митохондриялық тұқым қуалауға тән белгілер:
Белгілердің тіркес тұқым қуалауы.
Сызба – 2. Талдаушышағылыстыру №1.
Сызба – 3. Талдаушы шағылыстыру № 2
Нуклеин қышқылдары, құрылысы, жіктелуі
РНҚ –ның өмір сүруге қабілеттілігі:
Эукариоттардың генінің молеклалық құрылымы(схема түрінде)
Ген, классификациясы және функциясы
Геннің атқаратын қызметіне байлансыты гендер екі топқа бөлінеді:
Ағзалардың тіршілік қабілетіне байланысты гендер былай жіктеледі:
Нуклейн қышқылдары мен геннің медициналық маңызы
Сабақ 9 Ақпараттық дидактикалық блок
Репликацияның негізгі ферменттері:
Прокариотгендерініңактивтілігініңреттелуі:
Эукариоттар гендерінің экспрессиясының реттелуі
Гендерактивтілігініңгеномдықдеңгейде(ДНҚ)реттелуі
Эукариоттардың ДНҚ фракцияларының сипаттамалары
Кейбіргендердіңконститутивтік(тұрақты)экспрессиясы
Гендер активтілігінің транскрипциялық деңгейде реттелуі
Трансляциялықбақылау
Гендер экспрессиясын трансляциялық деңгейде бақылау прокариоттар мен эукариоттарда түрліше жолдармен іске асырылады. Прокариоттардың көпшілік а-РНҚ- ларының трансляциясы транскрипциямен қатар бір уақытта жүреді. Эукариоттық жасушаларда трансляция транскрипциядан кейін, процессинг нәтижесінде жетілген а-РНҚ түзілгеннен соң басталады.
Трансляциялық бақылаудың іске асырылу жолдары:
-
А-РНҚ-ның өмір сүру ұзақтығының жоғарылауы – информосомалардың немесе жасырын а-РНҚ-ның түзілуі арқылы іске асырылады. Мысалы, ұрықтанбаған жұмыртқа жасушасы әдетте биологиялық активті емес, бірақ ұрықтанудан соң іле-шала митотикалық аппараттың, жасушалық мембрананың белоктары мен гистондардың синтезі жүре бастайды. -
Белоктарсинтезделужылдамдығыныңреттелуі . Трансляциялық бақылаудың бұл механизмі де жетілген, ұрықтанбаған жұмыртқа жасушасында байқалады.Бұл жасушалар тыныштық күйде болып белок синтезделу процестері төменгі деңгейде жүріп жатады. Белок синтезделу қарқындылығының төмен болу себебі: а-РНҚ-ның жеткіліксіздігі емес, әзірше белгісіз, «жинақтаушы фактор» деп аталатын элементтің жетіспеуі. Бұл элемент рибосоманың а-РНҚ-мен қосылып кешен түзуіне қатынасатын болуы тиіс (рибосома+а- РНҚ). -
Мысалы, жібек көбелегінің (Bombyx mori) пілләсі түзілу үдерісінде «жібек» безі тек бір ғана белоктың түрін – фиброинды синтездейді. Фиброин белогының ұзақ уақыт синтезделуі, оның а-РНҚ-ның өмір сүруі ұзақтығының жоғарылауына байланысты.
Эукариоттардың қалыпты жағдайдағы а-РНҚ-сы 3 сағатқа жуық өмір сүреді де соңынан ыдырап кетеді. Ал фиброиндық а-РНҚ бірнеше күн өмір сүріп, сол уақыт аралығында фиброинның 109 молекуласын синтездеп үлгереді. «Жібек» безі жалпы алғанда 1015-ке жуық фиброин молекуласын (300 микрограмм) синтездейді. Фиброиндық а-РНҚ-ның өмір сүру мен қызмет атқаруын ұзартатын феномен болмаған жағдайда фиброинның қажетті мөлшері синтезделу үшін жібек көбелегінің құртына 25 есе артық ген қажет болып, олар 100 күндей жұмыс істеген болар еді.
Белоктардыңпосттрансляциялықмодификациялары
Кейбір белоктар синтез аяқталғаннан кейін құрылымдық өзгерістерге ұшырайды. Әдетте бірқатар белоктарда ыдырау немесе тримминг байқалады. Мысалы, инсулиннің (қызмет атқаруы) активтілігі геннің бірінші реттік өнімі ретінде синтезделген проинсулиннің орталық бөлігін ферменттер көмегімен алып тастау арқылы қамтамасыз етіледі. Көпшілік белоктардың активтілігі олардың басқа, құрылысы ұқсас немесе ұқсамайтын әртүрлі белоктармен байланысқан жағдайда іске асырылады. Мысалы, қызмет атқаруға қабілетті гемоглобин гемнен және глобиндік тізбектерден тұрады. Бұл байланыс бұзылса ағза темір жетіспейтін анемия ауруына шалдығады.
Гендерактивтілігініңгормональдықреттелуі
Жоғары сатыдағы өсімдіктер мен жануарларда жасушааралық өзара әрекеттесулер
олардың тіршілік үдерісінде маңызды роль атқарады. Жасушалардың өзара әрекеттесуі гормональдық әсер етуге байланысты іске асырылады. Гормондар тікелей жасуша – нысанаға немесе ұлпа – нысанаға әсер етеді. Пептидтік гормондар (инсулин эпинефрин т.б.) өздерінің ірі молекулалы болуына байланысты жасуша ішіне ене алмайды. Олар жасушаға рецепторлық белоктармен әрекеттесу арқылы әсер етеді. Рецепторлық белоктар әдетте нысаналық жасушаның мембраналарында орналасады. Осылай әсер ету арқылы пептидтік гормондар жасушаішілік циклдың АМФ (цАМФ)-тың деңгейін реттейді.
Циклдық АМФ протеин-киназа ферментін активтендіреді. Ал протеин-киназа ферменті көптеген арнайы қызмет атқаратын ферменттерді фосфорилдейді (активтендіріледі). Стероидтық гормондар (эстроген, прогестерон, тестерон, экдизон) жасуша ішіне оңай еніп, жасуша-нысана цитоплазмасындағы арнайы рецепторлық белоктармен берік байланысады. Түзілген
гормон + рецепторлық белок кешені арнайы гендердің транскрипциясын активтендіреді. Оның екі жолы бар: 1. Гормон + рецепторлық белок кешені арнайы гистонды емес белоктармен өзара әрекеттесіп нақты гендердің транскрипциясын активтендіреді. 2. Гормон + рецепторлық белок кешені гендердің энхансерлік және промоторлық
учаскелерінде орналасқан арнайы нуклеотидтер жүйесімен байланысып сол гендердің транскрипциясын активтендіреді (R.M. Evans, 1988). Осы
екі жағдайда да гормон + протеиндік кешен транскрипция үдерісіне прокариоттардағы САР-с АМР-кешенінің әсеріне ұқсас, позитивтік реттеуші әсер көрсетеді.
Олай болса, гендер активтілігі жасушалардың (ағзалардың) нақты тіршілік жағдайларына байланысты болып, тұқым қуалау ақпаратының жүзеге асырылуының барлық кезеңдерінде түрлі механизмдермен бақыланады.
Сабақ 12
Жасушалық цикл кезеңдерінің ретімен алмасып отыруы циклин және циклин-тәуелді киназа (Ц-ЦТК) кешендері, белок-ферменттерінің әсері арқылы іске асырылады.
Циклиннің ЦТК-ға жалғануы ЦТК-ны ғана активтендіріп қоймай, басқа да белоктарға әсері етеді. Жасшадағы циклиннің концентрациясы жасушалық цикл барысында циклды түрде өзгеріп отырады.
Жасушалық цикл ЦТК (Cdk) активтілігінің өзгеріуімен оның реттеуші суббірлігі болып табылатын циклиннің циклға сай түзіліп және қайта ыдырап отыруымен бақыланады. Түрлі циклиндердің синтезделіп және қайта ыдырап отыруы жасушалық циклдың фазаларының ретімен алмасып отыруын қамтамасыз етеді. Жасушалық циклдың түрлі фазаларында әртүрлі циклиндер түзіледі де ЦТК-мен жалғанып Ц-ЦТК кешендер түзеді. Бұл кешенде циклин активтендіруші , ал ЦТК -лар катализаторлар қызметін атқарады (сурет 1).
Олай болса төменде көрсетілген үдерістер жүргенде ғана митоздық циклдың реттелуінің бақылануы іске асырылады:
1. циклинге тәуелді киназа (Cdk) тек өзіне тән циклинмен қосылғанда ғана әсерін көрсетеді
2. Cdk фосфорилденгенде және дефосфорилденгенде
3.INK4 және KIP1 тұқымдасындағы арнайы ингибиторлық белоктармен әсерлескенде.
Сонымен Ц-ЦТК кешендерінің механизмдері жалпы белгілі бір ретпен жүретін төмендегі үдерістерді реттейді:
а) митоздық циклдың алдыңғы сатысын реттеуші факторлардың активсізденуін;
б) митоздық циклдың осы кезеңін қамтамасыз ететін факторлардың активтенуін;
в) жасуша бөлінуінің келесі кезеңіне қажетті факторлар кешенінің түзілуімен активтенуін.
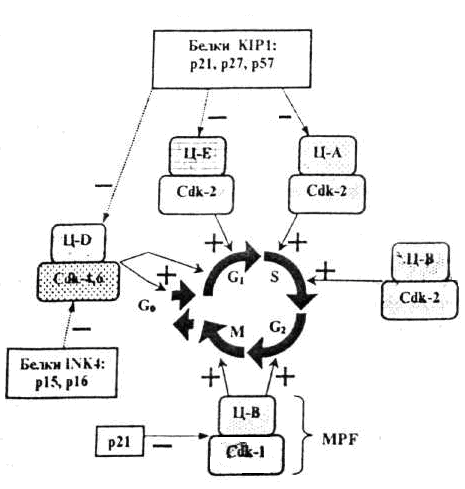
С. 1. Жасушалық циклдың түрлі фазаларын анықтайтын циклин- Cdk кешені
Ц- циклиндер, Cdk – циклин-тәуелді киназалар,
MPF –митозстимулдаушы фактор.
Төртбұрыш қоршауларда – ингибиторлар кешені.
Жасушаның бөлінуінің басталуы үшін сырқы орта жағдайдың қолайлы болуымен қатар арнайы сигналдық молекулалар – өсу факторлары (митогендер) және жасушаның қандайда бір беткейге бекінуі (эпителиоциттердің базальдық мембранаға, фибробластардың коллаген талшықтарына жалғануы) қажет және жасуша беткейіндегі рецепторлармен байланысқанда және контакталық тежелу болмаған жағдайда митоздық цикл механизмі іске қосылады.
Жасушалық циклді реттеудің сигналдық жолдары
Митотикалық цикл үдерісінің іске қосылуы циклин Д-ЦТК4 және/немесе циклин Д-ЦТК-6 кешенінің әсеріне байланысты болады. Бастаушы кешенді белсендіру үшін жасушадан тыс реттегіштер – өсу факторынан, яғни өсудің эпидермалдық факторынан (ЭФР) және антиген саналатын жасушадан (АПК) сигналдардың түсуі қажет.
Тирозинкиназа рецепторымен қабылданатын сигналдар каскадтық жолмен киназалық домен рецепторларын, митогенді активтендіреті протеинкиназаларды (МАПК) белсендіреді, олар митогеннің әсеріне тез әрі ерте жауап беретін (FOS, JUN) гендері үшін транскрипциялық факторлар (ТФ) болып табылады.
Ерте жауап беретін (FOS, JUN) гендерінің өнімдері өз кезегінде баяу жауап гендері үшін ТФ болады және оларды белсендіреді. Баяу жауап беретін ген өнімдері, оның ішінде ақуыз-ферменттер, циклин Д-ЦТК 4, ЦТК 6, митоздық циклдың G1–кезеңінің бірінші жартысында әсер ететін циклин-ЦТК кешендерінің пайда болуына әкеледі.
Сонымен қатар басқа да гендер, оның ішінде ақуыз Мус геніде белсенді болады. Мус ақуызы р27 генінің белсенділігін тежейді, р27 ақуызы циклин-ЦСК кешендерін тежейді және Сdc25a генін белсендіреді, оның өнімі спецификалық фосфатаза киназа мен ЦЗК2 фосфорилдеу арқылы оларды белсендіреді. Бұл үдерістердің нәтижесі әкеледі: