ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 05.12.2023
Просмотров: 3499
Скачиваний: 2
СОДЕРЖАНИЕ
Наиболее широко в ЦНС распространены медиаторы - амины:
Другие производные аминокислот - ГАМК, глицин, глютамин и др.
Название рецептора определено медиатором, с которым он взаимодействует:
Взаимодействие гормонов и парагормонов с клетками-мишенями
Понятие высших психических функций (Выготский)
Физиология газообмена в легких
Гуморальная, рефлекторная, нервная регуляция деятельности сердца
1.Общие свойства возбудимых тканей. Процесс возбуждения. Особенности местного и распространяющегося
Механизм формирования ПС связан с:
4. Современные представления о процессе возбуждения. Потенциал действия, его фазы.
8. Механизмы проведения возбуждения по нервным волокнам. Факторы, влияющие на скорость проведения
Механизмы проведения возбуждения по безмиелиновых нервным волокнам такой.
Закономерности проведения возбуждения через нервно-мышечный синапс:
В зависимости от частоты стимуляции выделяют следующие виды мышечного сокращения:
Тетанические сокращения отличается от одиночного следующими параметрами:
12. Функциональная характеристика гладких мышц.
13. Сила и работа мышц. Утомление и его особенности в целостном организме.
14. Нейрон как структурная и функциональная единица ЦНС. Его свойства и функции.
17-18. Возбуждение в ЦНС. Механизмы и закономерности передачи возбуждения в центральных
Особенности передачи возбуждения через центральные аксо-соматические химические синапсы.
Постсинаптическое гиперполяризацийне торможения.
Пресинаптическое деполяризации торможения.
Особенности передачи возбуждения в ЦНС:
Рефлекторная дуга имеет следующие звенья:
24. Рефлекс как элементарный акт нервной регуляции. Строение рефлекторной дуги
По расположению рецепторы подразделяют на:
По виду адекватного раздражителя, воспринимают рецепторы, их подразделяют на:
Физиологические механизмы кодирования информации в рецепторах.
26. Механизм кодирования информации в рецепторах. Адаптация рецепторов.
27. Общие принципы координационной деятельности ЦНС.
28. Суммация возбуждения, торможение нейронами ЦНС. Виды суммации и их значение
В зависимости от локализации рецепторного звена и эффекторного органа рефлексы делят на висцеро-
34. Сегментарные и надсегментарные центры вегетативной нервной системы
35. Гуморальная регуляция, её отличие от нервной. Факторы гуморальной регуляции.
Факторы гуморальной регуляции:
Механизм действия на клетки жирорастворимых гормонов:
Механизм действия жирорастворимых гормонов определяет следующие их особенности:
При воздействии на клетки-мишени водорастворимых гормонов образуются внутриклеточные посредники:
Классификация условных и безусловных рефлексов
- постоянство внутренней среды организма;
Современные представления о путях замыкания временных связей:
Эмоции выполнѐят две функции : сигнальную и регуляторную.
Эмоции делят на низшие и высшие.
Структурное обеспечение эмоций. Эмоциогенные структуры мозга.
Две сигнальные системы действительности
Типы высшей нервной деятельности
Общая характеристика восприятия
Безазотистые органические компоненты крови
Основные физико-химические константы крови:
Противосвертывающая система крови.
В норме гемоглобин содержится в виде нескольких соединений:
Методы исследования вентиляции легких:
Кислородная емкость крови, анализ кривой диссоциации:
Анализ кривой диссоциации НbО2:
^ Рефлекторная регуляция дыхания
второго порядка. Центр второго порядка может вырабатывать 40 - 60 импульсов в минуту.
^ Внутрисердечные механизмы регуляции.
Капиллярный кровоток и его особенности. Микроциркуляция и ее роль в механизме обмена жидкости и
Рефлекторная регуляция сердечно-сосудистой системы в зависимости от изменения положения тела в
Механизмы клубочковой фильтрации. Фильтрационное давление и факторы его определяющего. Состав
Механизм поддержания почками постоянства внутренней среды организма : рН, осмотического давления,
97. Функциональная система питания и пищеварения, ее основные звенья. Сенсорное насыщение. Функции
Пищеварение в полости рта. Состав и физиологическая роль слюны. Слюноотделение, его регуляция
101. Физиологическая роль печени, участие желчи в пищеварении. Факторы стимулирующие секрецию желчи,
106. Физиология щитовидной и околощитовидной желез
107. Физиология надпочечников. Роль гормонов коры и мозгового вещества в регуляции функции организма
Физиологическая характеристика обонятельной сенсорной системы. Механизмы восприятия запахов
6. Слюна обеспечивает минерализацию зубов т.к. содержит фосфор и кальций. Т.е. выполняет трофическую функцию.
7. Экскреторная. Со слюной выделяется небольшое количество продуктов белкового обмена - мочевина, мочевая кислота, креатинин, а также соли тяжелых металлов.
ЭКЗАМЕНАЦИОННЫЙ БИЛЕТ № 12
-
Механизм распространения возбуждения по миелинизированным и немиелинизированным волокнам. Законы проведения возбуждения по нерву. Физиологические основы методов местного обезболивания в стоматологии (инфильтрационного и проводникового).
Функцию быстрой передачи возбуждения к нервной клетке и от нее выполняют ее отростки – дендриты и аксоны, т.е. нервные волокна. В зависимости от структуры их делят на мякотные, имеющие миелиновую оболочку, и безмякотные. Эта оболочка формируется шванновскими клетками, являющиеся видоизмененными глиальными клетками. Они содержат миелин, который в основном состоит из липидов. Он выполняет изолирующую и трофическую функции. Одна шванновская клетка образует оболочку на 1 мм нервного волокна. Участки, где оболочка прерывается, т.е. не покрыты миелином, называют перехватами Ранвье. Ширина перехвата 1 мкм.
Проведение возбуждения по нервам подчиняется следующим законам:
1. Закон анатомической и физиологической целостности нерва.. 2. Закон двустороннего проведения возбуждения. Оно распространяется в обе стороны от места раздражения. В организме чаще всего возбуждение по афферентным путям идет к нейрону, а по эфферентным – от нейрона. 3. Закон изолированного проведения. Возбуждение не передается с одного нервного волокна на другое волокно, входящее в состав этого же нервного ствола.
4. Закон бездекрементного проведения. Возбуждение проводится по нервам без декремента, т.е. без затухания. 5. Скорость проведения прямо пропорциональна диаметру нерва. Т.к. по мембране безмякотного волокна местные токи текут не прерываясь, то такое проведение называется непрерывным. При непрерывном проведении местные токи захватывают большую поверхность волокна, поэтому им требуется, длительное время для прохождения по участку волокна. В результате дальность и скорость проведения возбуждения по безмякотным волокнам небольшая.
В мякотных волокнах участки, покрытые миелином, обладают большим электрическим сопротивлением. Поэтому непрерывное проведение ПД невозможно. При генерации ПД местные токи текут лишь между соседними перехватами. По закону "все или ничего" возбуждается ближайший к аксонному холмику перехват Ранвье, затем соседний нижележащий перехват и т.д. Такое проведение называется
сальтаторным (прыжком). При этом механизме ослабления местных токов не происходит и нервные импульсы распространяются на большое расстояние и с большой скоростью.
-
Артериальное давление как одна из физиологических констант организма. Строение и функция периферических и центральных звеньев системы саморегуляции артериального давления (рецепторы, центры, эффекторы). Дуга барорецептивного рефлекса.
Артериальное давление является одним из важнейших показателей гемодинамики и наиболее часто исследуется в клинике. Максимальное давление, которое возникает в результате систолы, называется систолическим артериальным давлением, а минимальное значение в диастолу - диастолическим давлением. У молодого человека нормальное систолическое давление составляет 110-120, а диастолическое 70-80 мм рт.ст. Разность между систолическим и диастолическим называется пульсовым давлением. Пульсовое давление, при прочих равных условиях пропорционально количеству крови, выбрасываемому сердцем при каждой систоле.
. Артериолы обладают толстой мышечной стенкой, с помощью которой меняется их просвет, и они являются главным регулятором уровня общего артериального давления.
Поэтому системное артериальное давление (САД) является результирующей величиной сердечного выброса (СВ) и общего периферического сопротивления сосудов (ОПСС):
САД = СВ • ОПСС.
Давление в крупных ветвях аорты (собственно артериальное) определяется как: АД = Q • R.
объемная скорость кровотока (Q) и сопротивление (R) сосудов.
регуляция осуществляется с помощью специального нервнорефлекторного механизма, называемого барорецептивным (прессорецептивным) рефлексом, информационная часть которого представлена собственными сосудистыми барорецепторами, расположенными в дуге аорты и в каротидном синусе.
Рецепторы реагируют на степень растяжения стенки сосуда пульсовыми или нарастающими колебаниями кровяного давления. В ответ на каждый систолический скачок давления барорецепторы генерируют залп импульсов, который затухает при диастолическом снижении давления. Чем выше давление крови в этих сосудах, тем сильнее раздражаются барорецепторы, и частота импульсов, посылаемых в сосудодвигательный центр, возрастает, и наоборот. От каротидного синуса в продолговатый мозг импульсы идут по чувствительному синокаротидному нерву (нерв Геринга), а от дуги аорты - по аортальному нерву, он же депрессорный нерв (нерв Циона-Людвига).
Импульсы направляются в вазомоторный центр, расположенный на дне 4-го желудочка продолговатого мозга.
сосудодвигательный центр располагается в продолговатом мозге и находится в состоянии тонической активности. В дальнейшем было установлено, что вазомоторный центр состоит из прессорной, депрессорной и кардиоингибирующей зон.
Прессорный отдел находится в состоянии постоянного возбуждения и посылает импульсы к периферическим сосудам через симпатические центры боковых рогов грудных сегментов спинного мозга и периферические симпатические нервы. Увеличение активности прессорного отдела вызывает повышение периферического сосудистого тонуса и увеличение системного артериального давления. Уменьшение его активности вызывает расширение сосудов и снижение давления.
Депрессорный отдел является центром, куда поступают импульсы непосредственно от сосудистых барорецепторов, под влиянием которых возрастает его активность. Влияние на артериальное давление он может оказывать только, угнетая активность прессорного отдела через тормозные вставочные интернейроны, что приводит к расширению сосудов и снижению артериального давления.
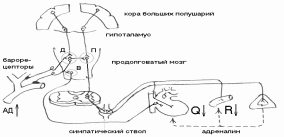
депрессорный отдел связан с кардиоингибирующим центром продолговатого мозга, представленным вегетативным ядром блуждающего нерва (на рис.23 – В). Импульсы, идущие от барорецепторов, одновременно с депрессорным центром повышают активность и центра вагуса, что приводит к урежению ритма сердца, уменьшению его выброса и снижению общего артериального давления
-
Вкусовой анализатор: механизм восприятия вкуса, проводниковый и корковый отделы анализатора. Методы исследования вкусового анализатора.
Значение вкусового анализатора заключается в апробации пищи при непосредственном соприкосновении ее со слизистой оболочкой полости рта.
Вкусовые рецепторы (периферический отдел) заложены в эпителии слизистой оболочки ротовой полости. Нервные импульсы по проводниковому пути, главным образом блуждающему, лицевому и языкоглоточному нервам, поступают в мозговой конец анализатора, располагающегося в ближайшем соседстве с корковым отделом обонятельного анализатора.
Вкусовые почки (рецепторы) сосредоточены, в основном, на сосочках языка. Больше всего вкусовых рецепторов имеется на кончике, краях и в задней части языка. Рецепторы вкуса располагаются также на задней стенке глотки, мягком небе, миндалинах, надгортаннике.
Раздражение одних сосочков вызывает ощущение только сладкого вкуса, других — только горького и т. д. Вместе с тем имеются сосочки, возбуждение которых сопровождается двумя или тремя вкусовыми ощущениями.
Основные вкусовые ощущения человека. У человека выявлено четыре четко различимых вкусовых ощущения: сладкое, кислое, соленое и горькое. Эти ощущения связаны со специфической чувствительностью различных участков поверхности языка. Вкус горького ощущается в первую очередь основанием языка, а сладкого — преимущественно кончиком языка. Ощущение кислого и соленого возникает при воздействии вкусовых раздражителей на боковые поверхности языка.
ЭКЗАМЕНАЦИОННЫЙ БИЛЕТ № 13
-
Роль И.П. Павлова в создании учения о пищеварении. Значение метода хронического эксперимента в изучении функций пищеварительных желез. Основные операции, разработанные в лаборатории Павлова.
Принцип метода заключается в хирургическ.подготовке животных, в ходе которой накладывают фистулу( отверстие снабженное специальной трубкой, выходящее наружу) того или иного отдела пищевар.тракта или выводных протоков пищеварит желез опыт ставится на выздоровевшем после операции животном. В лаборатории он успешно провел операцию ЭЗОФАГОТОМИИ( перерезка пищевода) после заживления раны проводил ,,мнимое кормление,, она ела но пища выпадала из отверстия пищевода , а из открытой желудочной фистулы изливался сок. Он в чистом виде получается у собак с изолированными, выкроенными в хир операциях из различных частей желудка желудочками.
-
Молекулярный механизм мышечного сокращения. Роль сократительных и регуляторных белков, ионов кальция и АТФ.
длина нитей актина и миозина в момент соскращения не изменяется. Поэтому Хаксли и Хэнсон разработали теорию скольжения нитей. Согласно этой теории мышца укорачивается в результате движения тонких актиновых нитей в промежутки между миозиновыми. Это приводит к укорочению каждого саркомера, образующего миофибриллы. Скольжение же нитей обусловлено тем, что при переходе в активное состояние головки отростков миозина связываются с центрами актиновых нитей и вызывают их движение относительно себя (гребковые движения). Но это последний этап всего сократительного механизма.
Сокращение начинается с того, что в области концевой пластинки двигательного нерва возникает ПД. Он с большой скоростью распространяется по сарколемме и переходит с неё по, системе поперечных трубочек СР, на продольные трубочки и цистерны. Возникает деполяризация мембраны цистерн и из них в саркоплазму высвобождаются ионы кальция. На нитях актина расположены молекулы еще двух белков –
тропонина и тропомиозина. При низкой (менее 10-8 моль) концентрации кальция, т.е. в состоянии покоя, тропомиозин блокирует присоединение мостиков миозина к нитям актина. Когда ионы кальция начинают выходить из СР, молекула тропонина изменяет свою форму таким образом, что освобождает активные центры актина от тропомиозина. К этим центрам присоединяются головки миозина и начинается скольжение за счет ритмического прикрепления и разъединения поперечных мостиков с нитями актина. При этом головки ритмически продвигаются по нитям актина к Z-мембранам. Для полного сокращения мышцы необходимо 50 таких циклов.
Передача сигнала от возбужденной мембраны к миофибриллам называется электромеханическим сопряжением. Когда генерация ПД прекращается и мембранный потенциал возвращается к исходному уровню, начинает работать Са-насос (фермент Са-АТФаза). Ионы кальция вновь закачиваются в цистерны саркоплазматического ретикулума и их концентрация падает ниже 10-8 моль. Молекулы тропонина приобретают исходную форму и тропомиозин вновь начинает блокировать активные центры актина. Головки миозина отсоединяются от них и мышца за счет эластичности приходит в исходное расслабленное состояние.
-
Жевательные и мимические мышцы, их участие в функции жевания, глотания, дыхания и речеобразования.
ЭКЗАМЕНАЦИОННЫЙ БИЛЕТ № 14
-
Потенциал действия кардиомиоцитов желудочков: фазы, ионный механизм, графическое изображение.
Возбуждение кардиомиоцита начинается быстрой деполяризацией его мембраны до нулевого уровн, которая продолжается быстрой сменой знака с – на +. Этим быстрым изменением полярности мембраны кардиомиоцита с уровня потенциала покоя начинается потенциал действия кардиомиоцита. Далее следует фаза реполяризации, она отличается от других возбудимых клеток.
Сначала реполяризация происходит быстро,затем медленно,на кривой записи потенциала действия кардиомиоцита видно плато. Затем осуществляется плавный переход к быстрой реполяризации мембраны до потенциала покоя. Длительность ПД намного выше чем у других клеток.
Эта особенность неспецифического эффекта нужна для его сокращения.
Потенциал покоя кардиомиоцитов варьирует в пределах от -50 до -95 мВ и обусловлен распределением ионов Na+, Ca2+, K+ и Cl- по обе стороны мембраны.
Наибольший вклад в значение потенциала вносит K+. В состоянии покоя мембрана обладает довольно высокой проницаемостью для ионов K.