ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 05.12.2023
Просмотров: 3292
Скачиваний: 2
СОДЕРЖАНИЕ
Наиболее широко в ЦНС распространены медиаторы - амины:
Другие производные аминокислот - ГАМК, глицин, глютамин и др.
Название рецептора определено медиатором, с которым он взаимодействует:
Взаимодействие гормонов и парагормонов с клетками-мишенями
Понятие высших психических функций (Выготский)
Физиология газообмена в легких
Гуморальная, рефлекторная, нервная регуляция деятельности сердца
1.Общие свойства возбудимых тканей. Процесс возбуждения. Особенности местного и распространяющегося
Механизм формирования ПС связан с:
4. Современные представления о процессе возбуждения. Потенциал действия, его фазы.
8. Механизмы проведения возбуждения по нервным волокнам. Факторы, влияющие на скорость проведения
Механизмы проведения возбуждения по безмиелиновых нервным волокнам такой.
Закономерности проведения возбуждения через нервно-мышечный синапс:
В зависимости от частоты стимуляции выделяют следующие виды мышечного сокращения:
Тетанические сокращения отличается от одиночного следующими параметрами:
12. Функциональная характеристика гладких мышц.
13. Сила и работа мышц. Утомление и его особенности в целостном организме.
14. Нейрон как структурная и функциональная единица ЦНС. Его свойства и функции.
17-18. Возбуждение в ЦНС. Механизмы и закономерности передачи возбуждения в центральных
Особенности передачи возбуждения через центральные аксо-соматические химические синапсы.
Постсинаптическое гиперполяризацийне торможения.
Пресинаптическое деполяризации торможения.
Особенности передачи возбуждения в ЦНС:
Рефлекторная дуга имеет следующие звенья:
24. Рефлекс как элементарный акт нервной регуляции. Строение рефлекторной дуги
По расположению рецепторы подразделяют на:
По виду адекватного раздражителя, воспринимают рецепторы, их подразделяют на:
Физиологические механизмы кодирования информации в рецепторах.
26. Механизм кодирования информации в рецепторах. Адаптация рецепторов.
27. Общие принципы координационной деятельности ЦНС.
28. Суммация возбуждения, торможение нейронами ЦНС. Виды суммации и их значение
В зависимости от локализации рецепторного звена и эффекторного органа рефлексы делят на висцеро-
34. Сегментарные и надсегментарные центры вегетативной нервной системы
35. Гуморальная регуляция, её отличие от нервной. Факторы гуморальной регуляции.
Факторы гуморальной регуляции:
Механизм действия на клетки жирорастворимых гормонов:
Механизм действия жирорастворимых гормонов определяет следующие их особенности:
При воздействии на клетки-мишени водорастворимых гормонов образуются внутриклеточные посредники:
Классификация условных и безусловных рефлексов
- постоянство внутренней среды организма;
Современные представления о путях замыкания временных связей:
Эмоции выполнѐят две функции : сигнальную и регуляторную.
Эмоции делят на низшие и высшие.
Структурное обеспечение эмоций. Эмоциогенные структуры мозга.
Две сигнальные системы действительности
Типы высшей нервной деятельности
Общая характеристика восприятия
Безазотистые органические компоненты крови
Основные физико-химические константы крови:
Противосвертывающая система крови.
В норме гемоглобин содержится в виде нескольких соединений:
Методы исследования вентиляции легких:
Кислородная емкость крови, анализ кривой диссоциации:
Анализ кривой диссоциации НbО2:
^ Рефлекторная регуляция дыхания
второго порядка. Центр второго порядка может вырабатывать 40 - 60 импульсов в минуту.
^ Внутрисердечные механизмы регуляции.
Капиллярный кровоток и его особенности. Микроциркуляция и ее роль в механизме обмена жидкости и
Рефлекторная регуляция сердечно-сосудистой системы в зависимости от изменения положения тела в
Механизмы клубочковой фильтрации. Фильтрационное давление и факторы его определяющего. Состав
Механизм поддержания почками постоянства внутренней среды организма : рН, осмотического давления,
97. Функциональная система питания и пищеварения, ее основные звенья. Сенсорное насыщение. Функции
Пищеварение в полости рта. Состав и физиологическая роль слюны. Слюноотделение, его регуляция
101. Физиологическая роль печени, участие желчи в пищеварении. Факторы стимулирующие секрецию желчи,
106. Физиология щитовидной и околощитовидной желез
107. Физиология надпочечников. Роль гормонов коры и мозгового вещества в регуляции функции организма
Физиологическая характеристика обонятельной сенсорной системы. Механизмы восприятия запахов
Остаточный объем (ОО) — объем воздуха, который остается в легких после максимального выдоха. ОО=1,0—1,5 л.
Легочные емкости:
- Жизненная емкость легких (ЖЕЛ) - максимальный объем воздуха, который можно выдохнуть после макс. вдоха. ЖЁЛ=ДО+РОвд+РОвыд.
У мужчин = 3,5—5,0 л и более. Уженщин = 3,0—4,0 л.
- Емкость вдоха (Евд)=ДО+РОвд. Евд= 2,0—2,3 л.
-Функциональная остаточная емкость (ФОЕ) — объем воздуха в легких после спокойного выдоха. ФОЕ=РОвыд+ОО= 1800-2500мл.
-Общая емкость легких (ОЕЛ) — объем воздуха в легких по окончании полного вдоха. ОЕЛ=ОО + ЖЕЛ, ОЕЛ=ФОЕ + Евд. у мужчин=6л, у женщин=5л.
Методы исследования вентиляции легких:
Измерение легочных объемов и емкостей имеет клиническое значение при исследовании функции легких у здоровых лиц и при диагностике заболевания легких человека. Измерение легочных объемов и емкостей обычно производят методами спирометрии, пневмотахометрии с интеграцией показателей, спирографиии
Отрицательное давление в плевральной полости (внутриплевральное) - ниже атмосферного в фазу выдоха на 6-8 см.вод.ст. а в экспирацию - на 4-5 см.вод.ст. Оно обусловлено эластической тягой легких, то есть, их постоянным стремлением спадаться, а также на отрицательное давление влияют активные силы, развиваемые дыхательными мышцами во время дыхательных движений. Расправленные легкие постоянно стремятся уменьшиться в объеме. Это обусловлено напряжением эластических волокон и поверхностным натяжением жидкости в альвеолах. Сила поверхностного натяжения тонкого слоя воды на поверхности альвеол всегда направлена на сжатие, и спадение альвеол Если сделать прокол грудной клетки и измерить внутри плевральное давление оно будет ниже атмосферного на 4-5 милм рт ст. Происхождение отрицательного давления в плевральной полости за счет неравномерного роста грудной клетки и легких легочная ткань отстает в росте от грудной клетки и поэтому легкие заполняясь воздухом старается спасьсся в результате эластической тяги и этим самым между париетальным и висцеральным листками образуется плевральная щель давление в которой ниже атмосферного. При повреждении грудной клетки или легких в плевральную полость может поступать воздух. При этом легкое спадается, попадание воздуха в плевральную полость называется пневмотораксом. Пневмоторакс может бать открытым, закрытым, клапанным односторонним и двусторонним двусторонний приводит к летальному исходу (объяснить меры помощи ) иногда роводят пневмоторакс с лечебной целью. При операции на легких применяется искусственная вентиляция легких. Дыхание у новорожденного легкие плода не сделавшего первого вдоха проверяют (объяснить). Легочная вентиляция определяется глубиной и частотой дыхания (16-20 в мин.). Легочные объемы.
69.Газообмен в легких и тканях. Парциальное давление газов (СО2, О2) в альвеолярном воздухе и напряжение газов в крои, тканевой жидкости и в клетках.
Физиология газообмена в легких
Физиологические основы газообмена в легких:
Газообмен осуществляется за счет диффузии газов через аэрогематический барьер.
1 этап: перенос газов по концентрационному градиенту через аэрогематический барьер,
2 этап: связывание газов в крови легочных капилляров.
Закон Фика: Qгаза= S*ДК*∆ P/ Т.
Qгаза - объем газа, проходящего через ткань в единицу времени.
S - площадь ткани,
ДК - диффузный коэффициент газа,
∆ Р - градиент парциального давления газа.
Т - толщина аэрогематического барьера.
Аэрогематический барьер: сурфактант - эпителий альвеол - интерстиция - эндотелий капилляров - плазма - эритроцит.
альвеола: рО2 = 40, рСО2=46;
венула: рО2 = 100, рСО2=40.
дельтаР О2 = 60, дельтаР СО2 = 6.
Поступление СО2 в легких из крови в альвеолы обеспечивается из следующих источников: 1) из СО2, растворенного в плазме крови (5—10%); 2) из гидрокарбонатов (80—90%); 3) из карбаминовых соединений эритроцитов (5—15%), которые способны диссоциировать.
Транспорт О2 и СО2 кровью:
Кислород в крови находится в растворенном виде и в соединении с гемоглобином. В плазме растворено очень небольшое количество кислорода. Поскольку растворимость кислорода при 37 °С составляет 0.225 мл * л-1 * кПа-1 (0.03 мл-л-1мм рт.ст.-1), то каждые 100 мл плазмы крови при напряжении кислорода 13.3 кПа (100 мм рг.ст.) могут переносить в растворенном состоянии лишь 0.3 мл кислорода. Этого недостаточно для жизнедеятельности организма. Отсюда ясна важность другого механизма переноса кислорода путем его соединения с гемоглобином.
Являясь конечным продуктом обмена веществ, СО2 находится в организме в растворенном и связанном состоянии. Коэффициент растворимости СО2составляет 0.231 ммольл-1 * кПа-1 (0.0308 ммольл-1 * мм рт.ст-1.), что почти в 20 раз выше, чем у кислорода. Однако, в растворенном виде переносится меньше 10% всего количества СО2 транспортируемого кровью. В основном, СО2переносится в химически связанном состоянии, главным образом, в виде бикарбонатов, а также в соединении с белками (так называемые карбоминовые, или карбосоединения).
Кислородная емкость крови, анализ кривой диссоциации:
Кислородная ёмкость крови - количество кислорода, которое может быть связано кровью при её полном насыщении; выражается в объёмных процентах. КЁК человека — около 18—20 об%.
Анализ кривой диссоциации НbО2:
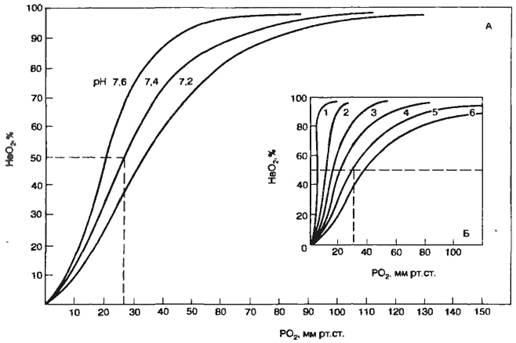
Зависимость степени оксигенации Нb от Рпарц. О2 в альвеолярном воздухе графически представляется в виде кривой диссоциации оксигемоглобина. Плато кривой диссоциации характерно для насыщенной О2 артериальной крови, а крутая нисходящая часть кривой — венозной крови в тканях.
Сродство Нb к О2 регулируется факторами метаболизма тканей: Ро2 pH, температурой и внутриклеточной концентрацией 2,3-дифосфоглицерата. сдвиг влево - легче идет насыщение О2: повышение рН, рО2, рСО2, понижение t, 2,3-ДФГ.
сдвиг вправо - легче идет отдача О2: понижение рН, рО2, рСО2, повышение 2,3-ДФГ, t.
Физиологические основы газообмена между кровью и тканями:
Обмен О2 между кровью капилляров и клетками тканей также осуществляется путем диффузии. Концентрационный градиент О2 между артериальной кровью (100 мм рт.ст.) и тканями (около 40 мм рт.ст) равен в среднем 60 мм рт.ст.
В ходе газообмена СО2 между тканями и кровью содержание НСОз- в эритроците повышается и они начинают диффундировать в кровь. Для поддержания электронейтральности в эритроциты начнут поступать из плазмы ионы С1- Наибольшее количество бикарбонатов плазмы крови образуется при участии карбоангидразы эритроцитов. Реакция СО2 с Нb приводит, во-первых, к высвобождению Н+; во-вторых, в ходе образования карбаминовых комплексов снижается сродство Нb к О2.
Кислород проникает из крови в клетки тканей путем диффузии, обусловленной разностью (градиентом) его парциальных давлений по обе стороны, так называемого гематопаренхиматоз-ного барьера. Так, среднее РО2 в артериальной крови составляет около 100 мм рт. ст., а в клетках, где кислород непрерывно утилизируется, стремится к нулю.кислород диффундирует в ткани не только из капилляров, но частично из артериол. Гематопаренхиматозный барьер помимо эндотелия кровеносного сосуда и клеточной мембраны включает и разделяющую их межклеточную (тканевую) жидкость. Перемещение тканевой жидкости, конвективные токи в ней могут способствовать транспорту кислорода между сосудом и клетками. Ту же роль, играют внутриклеточные цитоплазматические токи. И все же преобладающим механизмом переноса кислорода здесь служит диффузия, которая протекает тем интенсивнее, чем выше его потребление данной тканью.
Напряжение кислорода в тканях в среднем составляет 20−40 мм рт. ст. Функция газотранспортной системы организма в конечном счете направлена на поддержание парциального давления кислорода на клеточной Мембране не ниже критического, т.е. минимального, необходимого для работы ферментов дыхательной цепи в митохондриях. Для клеток, интенсивно потребляющих кислород, критическое РO2 составляет около 1 мм рт. ст. Напряжение O2 в тканях зависит не только от снабжения кислородом, но и от его потребления клетками. Наиболее чувствительны к недостатку кислорода клетки мозга, где окислительные процессы очень интенсивны.
В отличие от нервных клеток и клеток сердечной мышцы скелетные мышцы относительно устойчивы к кратковременному прекращению кислородного снабжения. Они используют при этом в качестве источника энергии анаэробный гликолиз. Кроме того, мышцы (особенно "красные") более выносливы к длительной работе, располагают незначительным резервом кислорода, запасенного в миоглобине. Миоглобин представляет собой дыхательный пигмент, подобный гемоглобину. Однако его сродство с кислородом значительно выше (P50 = 3 + 4 мм рт. ст.), поэтому он оксигенируется при относительно невысоком РO2 зато отдает кислород при очень низком его напряжении в тканях.
Перенос СО2 из клеток тканей в кровь тоже происходит главным образом путем диффузии, т.е. в силу разности напряжений СO2 по обе стороны гематопаренхиматозного барьера. Среднее артериальное значение РСО2 в среднем составляет 40 мм рт. ст., а в клетках может достигать 60 мм рт. ст. Локальное парциальное давление углекислого газа и, следовательно, скорости его диффузионного транспорта в значительной мере определяются продукцией СO2 (т.е. интенсивностью окисилительных процессов) в данном органе.
По той же причине РCO2 и PO2. B различных венах не одинаковы. Так, в крови, оттекающей от работающей мышцы, напряжение O2 гораздо ниже, а напряжение СO2 гораздо выше, чем, например, в крови, оттекающей от соединительной ткани. Поэтому для определения артериовенозной разницы, характеризующей суммарный обмен газов в организме, исследуют их содержание наряду с артериальной кровью (ее газовый состав практически одинаков в любой артерии) в смешанной венозной крови правого предсердия.
Коэффициент утилизации кислорода (КУО2) представляет собой часть кислорода, поглощаемую тканями из капиллярного русла; КУО2 определяют как отношение потребления кислорода к его доставке.В норме КУО2 = 24%.