ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 05.12.2023
Просмотров: 3461
Скачиваний: 2
СОДЕРЖАНИЕ
Наиболее широко в ЦНС распространены медиаторы - амины:
Другие производные аминокислот - ГАМК, глицин, глютамин и др.
Название рецептора определено медиатором, с которым он взаимодействует:
Взаимодействие гормонов и парагормонов с клетками-мишенями
Понятие высших психических функций (Выготский)
Физиология газообмена в легких
Гуморальная, рефлекторная, нервная регуляция деятельности сердца
1.Общие свойства возбудимых тканей. Процесс возбуждения. Особенности местного и распространяющегося
Механизм формирования ПС связан с:
4. Современные представления о процессе возбуждения. Потенциал действия, его фазы.
8. Механизмы проведения возбуждения по нервным волокнам. Факторы, влияющие на скорость проведения
Механизмы проведения возбуждения по безмиелиновых нервным волокнам такой.
Закономерности проведения возбуждения через нервно-мышечный синапс:
В зависимости от частоты стимуляции выделяют следующие виды мышечного сокращения:
Тетанические сокращения отличается от одиночного следующими параметрами:
12. Функциональная характеристика гладких мышц.
13. Сила и работа мышц. Утомление и его особенности в целостном организме.
14. Нейрон как структурная и функциональная единица ЦНС. Его свойства и функции.
17-18. Возбуждение в ЦНС. Механизмы и закономерности передачи возбуждения в центральных
Особенности передачи возбуждения через центральные аксо-соматические химические синапсы.
Постсинаптическое гиперполяризацийне торможения.
Пресинаптическое деполяризации торможения.
Особенности передачи возбуждения в ЦНС:
Рефлекторная дуга имеет следующие звенья:
24. Рефлекс как элементарный акт нервной регуляции. Строение рефлекторной дуги
По расположению рецепторы подразделяют на:
По виду адекватного раздражителя, воспринимают рецепторы, их подразделяют на:
Физиологические механизмы кодирования информации в рецепторах.
26. Механизм кодирования информации в рецепторах. Адаптация рецепторов.
27. Общие принципы координационной деятельности ЦНС.
28. Суммация возбуждения, торможение нейронами ЦНС. Виды суммации и их значение
В зависимости от локализации рецепторного звена и эффекторного органа рефлексы делят на висцеро-
34. Сегментарные и надсегментарные центры вегетативной нервной системы
35. Гуморальная регуляция, её отличие от нервной. Факторы гуморальной регуляции.
Факторы гуморальной регуляции:
Механизм действия на клетки жирорастворимых гормонов:
Механизм действия жирорастворимых гормонов определяет следующие их особенности:
При воздействии на клетки-мишени водорастворимых гормонов образуются внутриклеточные посредники:
Классификация условных и безусловных рефлексов
- постоянство внутренней среды организма;
Современные представления о путях замыкания временных связей:
Эмоции выполнѐят две функции : сигнальную и регуляторную.
Эмоции делят на низшие и высшие.
Структурное обеспечение эмоций. Эмоциогенные структуры мозга.
Две сигнальные системы действительности
Типы высшей нервной деятельности
Общая характеристика восприятия
Безазотистые органические компоненты крови
Основные физико-химические константы крови:
Противосвертывающая система крови.
В норме гемоглобин содержится в виде нескольких соединений:
Методы исследования вентиляции легких:
Кислородная емкость крови, анализ кривой диссоциации:
Анализ кривой диссоциации НbО2:
^ Рефлекторная регуляция дыхания
второго порядка. Центр второго порядка может вырабатывать 40 - 60 импульсов в минуту.
^ Внутрисердечные механизмы регуляции.
Капиллярный кровоток и его особенности. Микроциркуляция и ее роль в механизме обмена жидкости и
Рефлекторная регуляция сердечно-сосудистой системы в зависимости от изменения положения тела в
Механизмы клубочковой фильтрации. Фильтрационное давление и факторы его определяющего. Состав
Механизм поддержания почками постоянства внутренней среды организма : рН, осмотического давления,
97. Функциональная система питания и пищеварения, ее основные звенья. Сенсорное насыщение. Функции
Пищеварение в полости рта. Состав и физиологическая роль слюны. Слюноотделение, его регуляция
101. Физиологическая роль печени, участие желчи в пищеварении. Факторы стимулирующие секрецию желчи,
106. Физиология щитовидной и околощитовидной желез
107. Физиология надпочечников. Роль гормонов коры и мозгового вещества в регуляции функции организма
Физиологическая характеристика обонятельной сенсорной системы. Механизмы восприятия запахов
Возбудимые ткани
-
При действии на ткань электрического тока в ней исследовали уровень обменных процессов и деление клеток. Оказалось, что обмен веществ и деление клеток активизированы.Можно ли сделать вывод о том, что исследуемая ткань относится к возбудимым тканям?
-
Возбудимые ткани – это те,которые могут отвечать на раздражение возбуждение, т.е. давать активный ответ. -
Компоненты физиологического ответа могут быть специфическими (для мышечной ткани – сокращение, для нервной – формирование импульса, для железистой – секреция) и неспецифическими ( физиологические процессы,сопровождающиеся стуктурными,физико-химическими изменениями). -
Для всех возбудимых тканей характерно : повышение обмена веществ,теплопродукции и ИЗМЕНЕНИЕ электрического состояния клеточной мембраны. Исходя из этого, мы может сказать,что : данная ткань может быть возбудимой,так как при действии электрического тока в ней увеличился обмен веществ; мы можем отрицать,что эта ткань возбудима,тк в задаче не указано изменение электрического состояния мембраны,что есть важнейшим признаком возбудимой ткани.
-
Студент подготовил нервно-мышечный аппарат лягушки, но воспроизвести на нем второй опыт Гальвани не удается. В чем причина неудачи?
-
П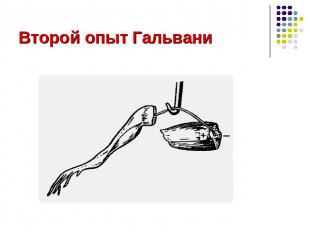 ри проведении второго опыта Гальвани с двумя нервно-мышечными препаратами ожидалось определить появление ПД,как доказательства существования собственного электрического тока в животных клетках.
ри проведении второго опыта Гальвани с двумя нервно-мышечными препаратами ожидалось определить появление ПД,как доказательства существования собственного электрического тока в животных клетках. -
Студент ожидал увидеть сокращение второго мышечного волокна. -
Механизм действия. При набрасывании нервного волокна первого препарата на второй так, чтоб оно касалось как поврежденного, так и неповрежденного участка второго препарата. За счет разности потенциалов между этими участками возникал электрический ток,который вызывает сокращение мышечного волокна первого препарата. -
Причиной неудачи могло стать нарушение проведения импульса по нервному волокну второго препарата, или же неправильное расположение нервного волокна на мышечном волокне первого препарата.
-
Нервное волокно поместили в раствор, в котором содержание отдельных ионов соответствовало их содержанию в тканевой жидкости. В этих условиях(контрольное исследование) был замерен МПП нервного волокна. Как расположены микроэлектроды? Каково значение МПП? Как изменится МПП при увеличении концентрации К+ в растворе в 50 раз?
-
МПП регестрируется внутриклеточным методом( один электрод на поверхности,другой – внутри). -
Величина МПП для нервного волокна приблизительно -50 - -80мВ -
При увеличении концентрации ионов Калия во внешней среде градиент концентрации Калия уменьшается,что приводит к ослаблению диффузного тока его из клетки и уменьшению МПП. -
При увеличении концентрации Калия в 50 раз вне клетки( с учетом того,что в клетке его(в норме) в 40 раз больше,чем вне ее) МПП исчезнет.
-
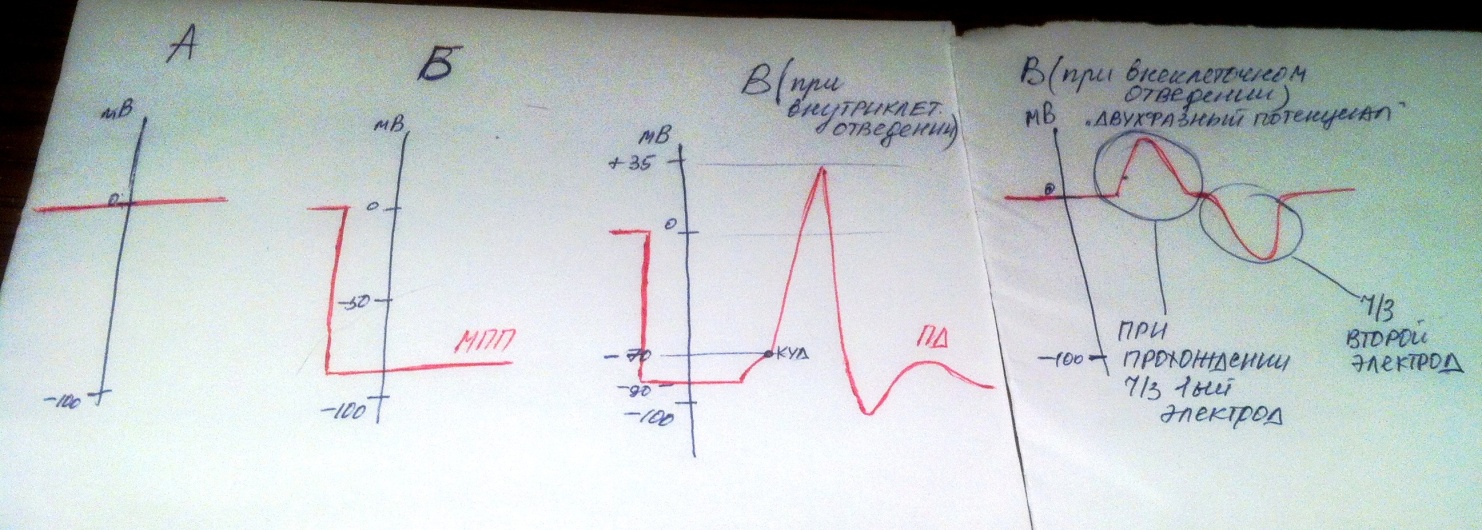
В эксперименте на возбужденной ткани разность потенциалов последовательно регестрировали: а)при расположении отводящих электродов на поверхности неповрежденной клетки, находящейся в покое; б) в момент проникновения электрода внутрь клетки; в)при действии на ткань порогового раздражения. Были ли зарегистрирована разность потенциалов во всех случаях? Опишите движение стрелки осциллографа. Когда был зарегистрирован моно- и дифазный ПД? Каковы условия их возникновения?
-
Разность потенциалов будет зарегестрирована в случае б и в, тк электроды расположены по обе стороны мембраны клетки(б) и имело место воздействие порогового раздражителя(в). В случае а она зарегестрирована не будет. -
В случае Б был зарегестрирован МПП, в случае В – ПД. -
Монофазный ПД зарегестрирован в случае Б и в случае В( если электроды при действии на ткань порогового раздражителя были расположены как при внутриклеточном отведении) -
Дифазный ПД(Возбуждение, которое представляет собой волну электроотрицательности, перемещаясь по мембране, доходит сначала до одного электрода, затем помещается между электродами, наконец достигает второго электрода, а затем распространяется дальше. ) будет зарегестрирован в случае В( если электроды расположены как при внеклеточном отведении). -
-
Известно,что фермент мембраны АТФ-аза с внутренней стороны мембраны связывается с Na+, с внешней – с К+. Объясните, почему при возбуждении усиливается работа Na+-K+- насоса.
-
АТФ-аза обеспечивает перенос 3 ионов Натрия из клетки в обмен на 2 иона Калия в клетку. -
Насос обеспечивает поддержание мембранного потенциала,клеточного объема, обменных процессов клетки. -
Работа насоса – активный процесс, тк он происходит с использованием энергии против градиента концентрации. -
Связь АТФ-азы и работы насоса: сам белок мембраны является АТФ-азой, только благодаря которой при гидролизе АТФ и присоединении фосфатной группы( а потом и дефосфорилирование при присоединении Калия) к белку-переносчику,может осуществляться присоединение ионов и их перенос. -
Работа насоса: 1. С внутренней стороны мембраны к молекуле белка-переносчика поступают АТФ и ионы натрия, а с наружной — ионы калия.2. Молекула переносчика осуществляет гидролиз одной молекулы АТФ.3. При участии трех ионов натрия за счет энергии АТФ к переносчику присоединяется остаток фосфорной кислоты (фосфорилирование переносчика); сами эти три иона натрия также присоединяются к переносчику.4. В результате присоединения остатка фосфорной кислоты происходит такое изменение формы молекулы переносчика (конформация), что ионы натрия оказываются по другую сторону мембраны, уже вне клетки.5. Три иона натрия выделяются во внешнюю среду, а вместо них с фосфорилированным переносчиком соединяются два иона калия.6. Присоединение двух ионов калия вызывает дефосфорилирование переносчика — отдачу им остатка фосфорной кислоты.7. Дефосфорилирование, в свою очередь, вызывает такую конформацию переносчика, что ионы калия оказываются по другую сторону мембраны, внутри клетки.8. Ионы калия высвобождаются внутри клетки, и весь процесс повторяется. -
При возбуждении(генерации ПД) роль насоса состоит в выкачивании ионов Натрия из клетки - в восстановлении МПП.
-
В эксперименте на нервном волокне блокада натриевых каналов мембраны ядом тетродотоксином изменила величину МПП. Что произошло с величиной МПП? Каков механизм действия?
-
При блокаде натриевых каналов( для величины МПП важны неспецифические каналы) прекращается диффузия Натрия в клетку по градиенту концентрации(уменьшается проницаемость мембраны для ионов Натрия). -
МПП зависит от ионной асимметрии и от работы механизма транспорта ионов,избирательной проницаемости мембраны. -
Для нервного волокна это имеет большое значение, тк генерация в нем МПП возможна только при закрытии большинства натриевых каналов и активности большинства калиевых. -
Потенциал покоя создаётся преимущественно за счёт выхода ионов калия по концентрационному градиенту из клетки. При этом натриевые каналы частично открыты, и некоторое количество ионов натрия проходит в клетку, уменьшая потенциал покоя. Следовательно, блокада натриевых каналов тетродоксином приведет к небольшому увеличению потенциала покоя.
-
Биопотенциалы нервной клетки регистрировали а)внеклеточным и б) внутриклеточным отведением. Было установлено, что поверхность клетки в покое электронейтральна(разность потенциалов отсутствует), величина МПП составляет -75мВ. Исследование повторили после отравления клетки препаратом, блокирующим процесс гликолиза. Объясните результаты измерения биопотенциалов отравленной клетки.
-
При нарушении процесса гликолиза происходит нарушение синтеза АТФ,что приводит к прекращению работы К-Na насоса. Соответственно прекращается поддержание МПП. Градиен концентрации ионов Натрия и Калия постепенно выравнивается, о чем говорит постепенное уменьшение МПП( что мы можем обнаружить при повторном исследовании). Электронейтральность наружной мембраны будет сохранятся.
-
При измерении ПД двух нервных волокон было установлено,что ПД первого больше ПД второго.При этом концентрация Натрия на поверхности обеих волокон одинакова, а внутри различна. Как зависит ПД от концентрации Натрия по обе стороны мембраны?
-
При рассмотрении изменения ПД мы учитываем изменения концентрация ионов Натрия потому,что процесс возбуждения,соответственно и генерация ПД, осуществляются благодаря току Натрия в клетку. -
Величина ПД тем больше,чем больше проницаемость мембраны для ионов Натрия.Чтобы повысить проницаемость,нужно увеличить градиент концентрации. Соответственно,если у первого волокна ПД меньше,чем у второго(при равной концентрации Натрия на поверхности мембраны), тем больше концентрация ионов Натрия у первого волокна,чем у второго, должна быть.
-
При раздражении мышечного волокна зарегистрировали дифазный ПД. Как было осущствлено исследование?
-
Чтобы получить дифазный ПД при раздражении волокна, нужно расположить оба электрода на неповрежденной поверхности волокна. Тогда: возбуждение, которое представляет собой волну электроотрицательности, перемещаясь по мембране, доходит сначала до одного электрода, затем помещается между электродами, наконец достигает второго электрода, а затем распространяется дальше. Что дает нам 2 фазы ПД. -
Чтобы получить монофазный ПД, нужно ввести 1 из электродов внутрь волокна.
-
ПД мышцы в условиях раздражения ее двигательного нерва был зарегестрирован при трех способах расположения регестрирующих электродов: 1) оба электрода на поверхности поврежденной мышци. 2) один электрод на неповрежденной поверхности, другой – внутри мышечного волокна. 3) один электрод на неповрежденном участке, другой – на поврежденном. Опишите особенности ПД в каждом случае.
-
В случае 1. Мы будем наблюдать дифазный ПД, что характерно для такого расположения электродов. В случае 2. И 3. Мы будем одинаково наблюдать монофазный ПД.Так как электрод,расположенный внутри мышечного волокна,будетидентичен в показаниях с электродом,расположенным на поврежденном участке мышечного волокна. -
Амплитуда ПД больше в опыте 2.,чем в опыте 1. ПОЧЕМУ ТАК.Я ОБЬЯСНИТЬ НЕ МОГУ
Вопрос №11
1) График ПД со следовыми потенциалами зарегистрированный в контрольном исследовании
0
-65
-90
мВ
t,c
деполяризация
реполяризация
следовая деполяризация
следовая гиперполяризация
КУД
МПП
овершут
2) В опыте 2 нарушен процесс деполяризации. Движущая сила — поступление ионов натрия, направленность — внутрь клетки.
3) В начале возбуждения внешние активационные ворота Na-каналов мембраны быстро открываются, проницаемость мембраны для натрия возрастает в несколько тысяч раз, натрий начинает лавинообразно поступать в клетку, обуславливаядеполяризацию. Для реполяризации характерно закрытие Na-каналов более медленными инактивационными воротами, расположенными на внутренней поверхности мембраны.
4) В опыте №2 (при блокировании натриевых каналов) деполяризация не будет развиваться в связи с тем, что натрий не поступает внутрь клетки. ПД не возникнет.
5) В опыте №3 изменяли концентрацию натрия в растворе, в который помещено нервное волокно. Это изменение концентрации подействовало аналогично блокаде Na-каналов, следовательно это изменение — резкое снижение концентрации натрия в растворе, либо полное изъятие натрия из раствора. В следствии этого натрий не поступает внутрь клетки, деполяризация не развивается, ПД отсутствует.
Вопрос №12
1) В нормальных условиях при возбуждении изменяется состояние Na-каналов мембраны. В начальной фазе потенциала действия - деполяризации Na-каналы быстро открываются. В начале фазы реполяризации Na-каналы закрываются.
2) В фазе деполяризации натрий лавинообразно поступает в клетку, заряжая внутреннюю поверхность мембраны положительно относительно внешней поверхности.
3) Блокаду натриевых каналов при введении тетрадотоксина можно определить в исследовании потенциала действия,который возникает (или не возникает) в ответ на стимуляцию клетки электрическим током.
4) Полное отсутствие потенциала действия при стимуляции клетки электрическим током свидетельствует о полной блокаде Na-каналов.
5) Эксперимент следует выполнять на нервной ткани или скелетной мышце, так как они относятся к возбудимым тканям и способны к генерации ПД.
Вопрос №13
1) При действии ТЭА (тетраэтиламмоний — блокирует калиевые каналы) возбудимость клетки сохраняется, так как начало процесса возбуждения клетки обусловлено поступлением натрия в клетку, и не связано с калием.
2) Во время фазы реполяризации ускоряется активация калиевых каналов, увеличивается диффузия калия наружу. Диффузия ионов калия во внеклеточную среду на фоне закрытия натриевых каналов приводит восстановлению потенциала покоя (реполяризация).
3) При действии ТЭА затрудняется выход ионов калия из цитоплазмы на наружную поверхность мембраны, поэтому удлиняется фаза реполяризации.
4) Механизм развития следовой гиперполяризации — остаточное усиление тока калия из клетки. Под действием ТЭА калиевые каналы блокируются, и следовая гиперполяризация не возникнет.
Вопрос №14
1) При нормальных условиях содержание ионов натрия во внеклеточной среде значительно превышает его содержание внутри клетки. У нервного волокна помещенного в бескислородную среду концентрация натрия по обе стороны мембраны выравнивается.
2) В бескислородной среде нарушается ресинтез АТФ и становится невозможной работа Na/K-насоса.
3) Градиент натрия создается при помощи активного транспорта - Na/K-насоса, который выводит из клетки 3 иона натрия в обмен на 2 иона калия.
4) Транспорт ионов с помощью Na/K-насоса относится к активным видам транспорта, так как осуществляется против градиента концентрации с затратой энергии АТФ.