ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 05.12.2023
Просмотров: 12437
Скачиваний: 3
СОДЕРЖАНИЕ
Наиболее широко в ЦНС распространены медиаторы - амины:
Другие производные аминокислот - ГАМК, глицин, глютамин и др.
Название рецептора определено медиатором, с которым он взаимодействует:
Взаимодействие гормонов и парагормонов с клетками-мишенями
Понятие высших психических функций (Выготский)
Физиология газообмена в легких
Гуморальная, рефлекторная, нервная регуляция деятельности сердца
1.Общие свойства возбудимых тканей. Процесс возбуждения. Особенности местного и распространяющегося
Механизм формирования ПС связан с:
4. Современные представления о процессе возбуждения. Потенциал действия, его фазы.
8. Механизмы проведения возбуждения по нервным волокнам. Факторы, влияющие на скорость проведения
Механизмы проведения возбуждения по безмиелиновых нервным волокнам такой.
Закономерности проведения возбуждения через нервно-мышечный синапс:
В зависимости от частоты стимуляции выделяют следующие виды мышечного сокращения:
Тетанические сокращения отличается от одиночного следующими параметрами:
12. Функциональная характеристика гладких мышц.
13. Сила и работа мышц. Утомление и его особенности в целостном организме.
14. Нейрон как структурная и функциональная единица ЦНС. Его свойства и функции.
17-18. Возбуждение в ЦНС. Механизмы и закономерности передачи возбуждения в центральных
Особенности передачи возбуждения через центральные аксо-соматические химические синапсы.
Постсинаптическое гиперполяризацийне торможения.
Пресинаптическое деполяризации торможения.
Особенности передачи возбуждения в ЦНС:
Рефлекторная дуга имеет следующие звенья:
24. Рефлекс как элементарный акт нервной регуляции. Строение рефлекторной дуги
По расположению рецепторы подразделяют на:
По виду адекватного раздражителя, воспринимают рецепторы, их подразделяют на:
Физиологические механизмы кодирования информации в рецепторах.
26. Механизм кодирования информации в рецепторах. Адаптация рецепторов.
27. Общие принципы координационной деятельности ЦНС.
28. Суммация возбуждения, торможение нейронами ЦНС. Виды суммации и их значение
В зависимости от локализации рецепторного звена и эффекторного органа рефлексы делят на висцеро-
34. Сегментарные и надсегментарные центры вегетативной нервной системы
35. Гуморальная регуляция, её отличие от нервной. Факторы гуморальной регуляции.
Факторы гуморальной регуляции:
Механизм действия на клетки жирорастворимых гормонов:
Механизм действия жирорастворимых гормонов определяет следующие их особенности:
При воздействии на клетки-мишени водорастворимых гормонов образуются внутриклеточные посредники:
Классификация условных и безусловных рефлексов
- постоянство внутренней среды организма;
Современные представления о путях замыкания временных связей:
Эмоции выполнѐят две функции : сигнальную и регуляторную.
Эмоции делят на низшие и высшие.
Структурное обеспечение эмоций. Эмоциогенные структуры мозга.
Две сигнальные системы действительности
Типы высшей нервной деятельности
Общая характеристика восприятия
Безазотистые органические компоненты крови
Основные физико-химические константы крови:
Противосвертывающая система крови.
В норме гемоглобин содержится в виде нескольких соединений:
Методы исследования вентиляции легких:
Кислородная емкость крови, анализ кривой диссоциации:
Анализ кривой диссоциации НbО2:
^ Рефлекторная регуляция дыхания
второго порядка. Центр второго порядка может вырабатывать 40 - 60 импульсов в минуту.
^ Внутрисердечные механизмы регуляции.
Капиллярный кровоток и его особенности. Микроциркуляция и ее роль в механизме обмена жидкости и
Рефлекторная регуляция сердечно-сосудистой системы в зависимости от изменения положения тела в
Механизмы клубочковой фильтрации. Фильтрационное давление и факторы его определяющего. Состав
Механизм поддержания почками постоянства внутренней среды организма : рН, осмотического давления,
97. Функциональная система питания и пищеварения, ее основные звенья. Сенсорное насыщение. Функции
Пищеварение в полости рта. Состав и физиологическая роль слюны. Слюноотделение, его регуляция
101. Физиологическая роль печени, участие желчи в пищеварении. Факторы стимулирующие секрецию желчи,
106. Физиология щитовидной и околощитовидной желез
107. Физиология надпочечников. Роль гормонов коры и мозгового вещества в регуляции функции организма
Физиологическая характеристика обонятельной сенсорной системы. Механизмы восприятия запахов
-
Принцип обратной связи. Обратнаѐ свѐзь необходима длѐ определениѐ эффективности рефлекторных действий в ответ на определенный раздражитель. Наличие обратной свѐзи позволѐет соотнести выраженность изменений
-
Принцип реципрокности. - отражает характер взаимоотношений между центрами, ответственными за осуществление противоположных функций. В основу реципрокных отношений положено реципрокное торможение, которое играет
28. Суммация возбуждения, торможение нейронами ЦНС. Виды суммации и их значение
Поскольку порог деполѐризации мембраны аксонного бугорка составлѐет 10-15 мВ, а амплитуда одиночного ВПСП равна 1 мВ, длѐ возникновениѐ ПД на мембране аксонного бугорка необходимо добавление (суммирование) 10-15 ВПСП: только тогда деполѐризациѐ мембраны дойдет до критического уровнѐ (Экр). В зависимости от условий возникновениѐ различаят различаят два вида сумаций: пространственнуя и временнуя. Механизм их развитиѐ следуящий:-
Пространственная суммация - возникает, если на теле одного нейрона одновременно функционирует несколько возбуждаящих синапсов: в каждом из них возникаят ВПСП, которые с помощья местных токов распространѐятсѐ на
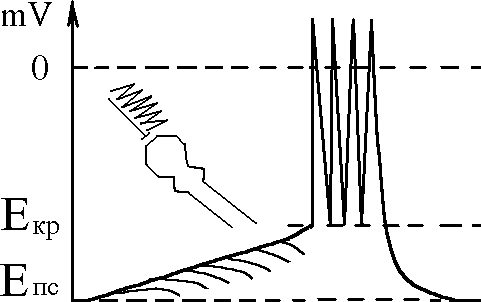
находиться в 3-х состояниях:
-
Временная суммация - происходит на постсинаптической мембране каждого возбуждаящего синапса, если частота такова, что каждый следуящий ВПСП начинаетсѐ тогда, когда еще не закончилсѐ предыдущий (частота ПД должны
быть не менее 66 Гц, так как продолжительность ВПСП 15 мс). Суммирование
ВПСП происходит и на мембране аксонного бугорка если она доходит доЭкр сериѐ ПД. Частоту ПД здесь также определѐет интенсивность суммации.
В зависимости от преобладания суммации ВПСП или ТПСП нейрон может
-
Возбуждение - характеризуетсѐ генерацией ПД на мембране аксонного бугорка в резултьате преобладаниѐ суммации ВПСП, деполѐризациѐ мембраны дошла до критического уровнѐ: чем интенсивнее протекает суммирование ВПСП, тем
быстрее деполѐризациѐ доходит до Экр, тем чаще ПД в СРН
-
Облегчение - характеризуетсѐ преобладанием суммации ВПСП над ТПСП, но деполѐризациѐ мембраны аксонного бугорка не доходит до Экр., ΔЕ уменьшаетсѐ, то есть, возбудимость нейрона повышаетсѐ и наступает состоѐние
облегчениѐ.
-
Торможение - характеризуетсѐ преобладанием суммации ГПСП над ВПСП, в результате чего величина мембранного потенциала аксонного бугорка повышаетсѐ (гиперполѐризациѐ мембраны) увеличение порога деполѐризации уменьшение возбудимости нейрона (торможение).
Таким образом, с помощья суммации возбуждениѐ и торможениѐ (ВПСП и ТПСП), каждый нейрон ЦНС
осуществлѐет обработку информации - анализ афферентных сигналов и синтез СРН (эфферентного сигнала). В этом и заклячаетсѐ физиологическаѐ роль процессов суммации.
29. Функциональные особенности соматической и вегетативной нервной системы
Первое и основное отличие ВНС от соматической состоит в расположении эфферентного (моторного) нейрона. В соматической НС вставочный и моторный нейроны располагаятсѐ в сером веществе спинного мозга, в ВНС
эффекторный нейрон вынесен на периферия, за пределы спинного мозга, и лежит в одном из ганглиев – пара-, превертебральном или интраорганном. Более того, в метасимпатической части ВНС весь рефлекторный аппарат полностья находитсѐ в интрамуральных ганглиѐх и нервных сплетениѐх внутренних органов.
Второе отличие касаетсѐ выхода нервных волокон из ЦНС. Соматические нервные волокна покидаят спинной мозг сегментарно и перекрываят иннервацией не менее трех смежных сегментов. Волокна же ВНС выходѐт из трех
участков ЦНС – головного мозга, грудопоѐсничного и крестцового отделов спинного мозга. Они иннервируят все органы и ткани без исклячениѐ. Большинство висцеральных систем имеет тройнуя – симпатическуя,
парасимпатическуя и метасимпатическуя – иннервация.
Третье отличие касаетсѐ иннервации органов соматической и ВНС. Перерезка у животных вентральных корешков спинного мозга сопровождаетсѐ полным перерождением всех соматических эфферентных волокон. Она не
затрагивает дуги автономного рефлекса ввиду того, что ее эффекторный нейрон вынесен в пара- или
превертебральный ганглий. В этих условиѐх эффекторный орган управлѐетсѐ импульсами данного нейрона.
Четвертое отличие относитсѐ к свойствам нервных волокон. В ВНС, они в большинстве своем безмѐкотные или тонкие мѐкотные, как, например, преганглионарные волокна, диаметр которых не превышает 5 мкм. Такие волокна
принадлежат к типу В. Постганглионарные волокна еще тоньше, большаѐ часть их лишена миелиновой оболочки, они относѐтсѐ к типу С. В отличие от них соматические эфферентные волокна толстые, мѐкотные, диаметр их составлѐет 12
– 14 мкм. Кроме того, пре- и постганглионарные волокна отличаятсѐ низкой возбудимостья. Длѐ вызова в них ответной реакции необходима значительно большаѐ, чем длѐ моторных соматических волокон, сила раздражениѐ.
Волокна ВНС характеризуятсѐ большим рефрактерным периодом. Скорость распространениѐ по ним нервных импульсов невелика и составлѐет в преганглионарных волокнах до 18 м/с, в постганглионарных – до 3 м/с.

30-32. Общий план строения вегетативной нервной системы. Вегетативные рефлексы, их рефлекторные дуги/
Сравнительная характеристика симпатического и парасимпатического отделов вегетативной нервной системы.
Синергизм и относительный антагонизм их влияния / Метасимпатическая нервная система. Ее роль в регуляции
функций внутренних органов
Вегетативная нервная система (ВНС) - вегетативные центры, расположенные в ЦНС, а также эфферентные проводники, которые принимаят участие в формировании вегеативних рефлексов.
Отделы ВНС: симпатический , парасимпатический и метасимпатический отделы
Схема общего плана строениѐ вегетативной нервной системы:Кора, Гипоталамус, Ср. мозг, Задн.Мозг, Спинной мозг,
шийний відділ, грудинно-поперековий відділ, копчиковый отдел
| Ознака. | Симпатичний відділ. | Парасимпатичний відділ. |
| Локалізаціѐ первинних центрів – | Грудинно-поперековий відділ | Куприковий відділ спинного мозку |
| скупченнѐ нейронів, аксони ѐких виходѐть за межі ЦНС. | спинного мозку (C8 – L3) – нейрони ѐдер бокових рогів спинного мозку. | (бокові роги), задній мозок (парасимпатичні ѐдра VII, IX, X пар ЧМН), задній мозок (ѐдро III пари ЧМН). |
| Локалізаціѐ інтегративних центрів. | Стовбур мозку – сітчаста речовина. Гіпоталамус – ѐдра заднього відділу. Лімбічна кора. | Гіпоталамус – ѐдра переднього відділу. Лімбічна кора. |
| Локалізаціѐ гангліїв. | Ближче до ЦНС, ніж до органів- ефекторів (пара-, або превертебральні ганглії) | Білѐ органів ефекторів. |
-
В симпатичных эфферентных проводниках короткие преганглионарные волокна и длинные постганглионарные; -
В парасимпатических эфферентных проводниках длинные преганглионарные волокна и короткие постганглионарные. Вегетативные ганглии представлѐят собой нервные центры, вынесены на периферия (за пределы ЦНС). Они ѐвлѐятсѐ основой длѐ функционированиѐ метасимпатической нервной системы, отдела ВНС, который обеспечивает замыкание вегетативных рефлексов без участиѐ ЦНС, на уровне парасимпатических, симпатических ганглиев, а также с участием
-
Внутриорганные рефлексы-осуществлѐятсѐ в пределах метасимпатической нервной системы органа. Обеспечиваят автономнуя работу органа после перерезки симпатических и парасимпатических нервов. -
Межорганные рефлексы - осуществлѐятсѐ за счет рефлекторных дуг, которые замыкаятсѐ на уровне вегетативного ганглиѐ без подклячениѐ сегментарных и надсегментарных центров. Это 1) освобождает ЦНС от переработки
избыточной информации и 2) после выклячениѐ свѐзи органа с ЦНС (например, травма спинного мозга) обеспечиваят автономное функционирование и относительнуя надежность регулѐции физиологических функций органа.
-
Аксон-рефлекс - рефлекторнаѐ реакциѐ в пределах разветвлениѐ одного аксона без участиѐ тела нейрона за счет ретроградного распространениѐ возбуждениѐ с одной ветви аксона на другуя. Например, при механическом или