ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 05.12.2023
Просмотров: 3490
Скачиваний: 2
СОДЕРЖАНИЕ
Наиболее широко в ЦНС распространены медиаторы - амины:
Другие производные аминокислот - ГАМК, глицин, глютамин и др.
Название рецептора определено медиатором, с которым он взаимодействует:
Взаимодействие гормонов и парагормонов с клетками-мишенями
Понятие высших психических функций (Выготский)
Физиология газообмена в легких
Гуморальная, рефлекторная, нервная регуляция деятельности сердца
1.Общие свойства возбудимых тканей. Процесс возбуждения. Особенности местного и распространяющегося
Механизм формирования ПС связан с:
4. Современные представления о процессе возбуждения. Потенциал действия, его фазы.
8. Механизмы проведения возбуждения по нервным волокнам. Факторы, влияющие на скорость проведения
Механизмы проведения возбуждения по безмиелиновых нервным волокнам такой.
Закономерности проведения возбуждения через нервно-мышечный синапс:
В зависимости от частоты стимуляции выделяют следующие виды мышечного сокращения:
Тетанические сокращения отличается от одиночного следующими параметрами:
12. Функциональная характеристика гладких мышц.
13. Сила и работа мышц. Утомление и его особенности в целостном организме.
14. Нейрон как структурная и функциональная единица ЦНС. Его свойства и функции.
17-18. Возбуждение в ЦНС. Механизмы и закономерности передачи возбуждения в центральных
Особенности передачи возбуждения через центральные аксо-соматические химические синапсы.
Постсинаптическое гиперполяризацийне торможения.
Пресинаптическое деполяризации торможения.
Особенности передачи возбуждения в ЦНС:
Рефлекторная дуга имеет следующие звенья:
24. Рефлекс как элементарный акт нервной регуляции. Строение рефлекторной дуги
По расположению рецепторы подразделяют на:
По виду адекватного раздражителя, воспринимают рецепторы, их подразделяют на:
Физиологические механизмы кодирования информации в рецепторах.
26. Механизм кодирования информации в рецепторах. Адаптация рецепторов.
27. Общие принципы координационной деятельности ЦНС.
28. Суммация возбуждения, торможение нейронами ЦНС. Виды суммации и их значение
В зависимости от локализации рецепторного звена и эффекторного органа рефлексы делят на висцеро-
34. Сегментарные и надсегментарные центры вегетативной нервной системы
35. Гуморальная регуляция, её отличие от нервной. Факторы гуморальной регуляции.
Факторы гуморальной регуляции:
Механизм действия на клетки жирорастворимых гормонов:
Механизм действия жирорастворимых гормонов определяет следующие их особенности:
При воздействии на клетки-мишени водорастворимых гормонов образуются внутриклеточные посредники:
Классификация условных и безусловных рефлексов
- постоянство внутренней среды организма;
Современные представления о путях замыкания временных связей:
Эмоции выполнѐят две функции : сигнальную и регуляторную.
Эмоции делят на низшие и высшие.
Структурное обеспечение эмоций. Эмоциогенные структуры мозга.
Две сигнальные системы действительности
Типы высшей нервной деятельности
Общая характеристика восприятия
Безазотистые органические компоненты крови
Основные физико-химические константы крови:
Противосвертывающая система крови.
В норме гемоглобин содержится в виде нескольких соединений:
Методы исследования вентиляции легких:
Кислородная емкость крови, анализ кривой диссоциации:
Анализ кривой диссоциации НbО2:
^ Рефлекторная регуляция дыхания
второго порядка. Центр второго порядка может вырабатывать 40 - 60 импульсов в минуту.
^ Внутрисердечные механизмы регуляции.
Капиллярный кровоток и его особенности. Микроциркуляция и ее роль в механизме обмена жидкости и
Рефлекторная регуляция сердечно-сосудистой системы в зависимости от изменения положения тела в
Механизмы клубочковой фильтрации. Фильтрационное давление и факторы его определяющего. Состав
Механизм поддержания почками постоянства внутренней среды организма : рН, осмотического давления,
97. Функциональная система питания и пищеварения, ее основные звенья. Сенсорное насыщение. Функции
Пищеварение в полости рта. Состав и физиологическая роль слюны. Слюноотделение, его регуляция
101. Физиологическая роль печени, участие желчи в пищеварении. Факторы стимулирующие секрецию желчи,
106. Физиология щитовидной и околощитовидной желез
107. Физиология надпочечников. Роль гормонов коры и мозгового вещества в регуляции функции организма
Физиологическая характеристика обонятельной сенсорной системы. Механизмы восприятия запахов
Как правило, подкрепление развертывается во временной динамике. Временная динамика подкрепления складывается в процессе нередко продолжительного достижения субъектами, испытывающими ту или иную потребность, часто многочисленных промежуточных и только после этого - конечного результатов, удовлетворяющих эту потребность. Одни результаты поведения при этом могут приближать субъекта к удовлетворению исходной потребности, иногда в какой-то степени удовлетворять ее. Другие результаты - наоборот, препятствуют удовлетворению субъектом доминирующей потребности. При взаимодействии субъекта с разнообразными промежуточными и конечным результатами при обучении происходит формирование и ооогащение аппарата акцептора результата действия путем построения широко разветвленных по структурам мозга динамических энграмм подкрепления.
51. Базальные ганглии, их афферентные и эфферентные связи. Роль базальных ядер в формировании мышечного тонуса и сложных двигательных актов. Последствия повреждения базальных ганглий и их связей.
Базальные ганглии, названные так гистологами прошлого века, представляют собой структуры ядерного типа, которые располагаются в толще белого вещества переднего мозга ближе к его основанию.
У млекопитающих к базальным ганглиям относятся сильно вытянутое в длину и изогнутое хвостатое ядро и заложенное в толще белого веществачечевицеобразное ядро. Двумя белыми пластинками оно подразделяется на три части: наиболее крупную, лежащую латерально скорлупу ибледный шар, состоящий из внутреннего и внешнего отделов.
Эти анатомические образования формируют так называемуюстриопаллидарную систему, которая по филогенетическим и функциональным критериям разделяется на древний палеостриатум и неостриатум. Палеостриатум представлен бледным шаром, анеостриатум, появляющийся впервые у рептилий, состоит из хвостатого ядра и скорлупы, которые объединяются под названием полосатого тела или стриатума. Хвостатое ядро и скорлупа связаны анатомически и характеризуются чередованием белого и серого вещества, что оправдывает возникновение термина полосатое тело.
К стриопаллидарной системе часто относят также субталамическое ядро (люисово тело) и черную субстанцию среднего мозга, которые образуют с базальными ганглиями функциональное единство. Полосатое тело состоит главным образом из мелких клеток, аксоны которых направляются к бледному шару и черной субстанция среднего мозга.
Полосатое тело является своеобразным коллектором афферентных входов, идущих к базальным ганглиям. Главными источниками этих входов служат новая кора (преимущественно сенсомоторная), неспецифические ядра таламуса и дофаминэргические пути от черной субстанции.
В противоположность полосатому телу бледный шар состоит из крупных нейронов и является сосредоточением выходных, эфферентных путей стриопаллидарной системы. Аксоны локализованных в бледном шаре нейронов подходят к различным ядрам промежуточного и среднего мозга, в том числе и к красному ядру, где начинается
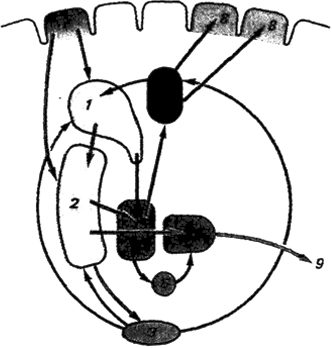
Рис. 3.15. Афферентные и эфферентные связи (шальных ганглиев: 1 - хвостатое ядро, 2 - скорлупа, 3 - черная субстанция, 4 - таламус, 5, 5' - внутренний и внешний отделы бледного шара, б - субталамическое ядро, 7 - сенсомоторная кора, 8 - двигательная кора, 9 - пути к стволовой части мозга; синими стрелками обозначены афферентные пути, красными - эфферентные, черными - межъядерные взаимодействия
руброспинальный тракт экстрапирамидной системы двигательной регуляции.
Другой важный эфферентный тракт идет от внутреннего отдела бледного шара к передневентральному и вентролатеральному ядрам таламуса, а оттуда продолжается к двигательным областям коры головного мозга. Наличие этого пути обусловливает многозвенную петлеобразную связь между сенсомоторными и двигательными областями коры, которая осуществляется через полосатое тело и бледный шар к таламусу. Примечательно, что в составе этого стриапаллидоталамокортикального пути базальные ганглии выполняют роль афферентного звена по отношению к моторным областям коры головного мозга. Многочисленные связи стриопаллидарной системы с различными отделами мозга свидетельствуют об ее участии в процессах интеграции, однако до настоящего времени в знании о функциях базальных ганглиев остается много невыясненного.
Базальные ганглии играют важную роль в регуляции движений и сенсомоторной координации. Известно, что при повреждении полосатого тела наблюдается атетоз - медленные червеобразные движения кистей и пальцев рук. Дегенерация клеток стриатума вызывает также другое заболевание - хорею, выражающуюся в судорожных подергиваниях мимических мышц и мускулатуры конечностей, которые наблюдаются в покое и при выполнении произвольных движений. Однако попытки выяснить этиологию этих нарушений в экспериментах на животных не дали результатов. Разрушение хвостатого ядра у собак и кошек не приводило к возникновению гиперкинезов, характерных для описанных выше заболеваний.
Локальное электрическое раздражение некоторых участков стриатума вызывает у животных так называемые циркуляторные двигательные реакции, характеризующиеся поворотом головы и
туловища в сторону, противоположную раздражению. Раздражение других участков полосатого тела, напротив, приводит к торможению двигательных реакций, вызванных различными сенсорными раздражениями.
Наличие определенных расхождений между данными эксперимента и клиники, по-видимому, свидетельствует о возникновении системных нарушений механизмов регуляции движений при патологических процессах в базальных ганглиях. Очевидно, эти нарушения связаны с изменениями функции не только полосатого тела, но и других структур. В качестве примера можно рассмотреть возможный патофизиологический механизм возникновения дрожательного паралича - синдрома Паркинсона. Этот синдром связан с повреждением базальных ганглиев и характеризуется комплексом таких симптомов, как акинезия - малая подвижность и затруднения при переходе от покоя к движению; восковидная ригидность, или гипертонус, не зависящий от положения суставов и фазы движения; статический тремор, наиболее выраженный в дистальных отделах конечностей.
Все эти симптомы, согласно современным представлениям, обусловлены гиперактивностью базальных ганглиев, которая возникает при повреждении дофаминэргического (по всей вероятности, тормозного) пути, который идет от черной субстанции к полосатому телу. Таким образом, этиология синдрома Паркинсона обусловлена дисфункцией как полосатого тела, так и структур среднего мозга, которые функционально объединены в стриопаллидарную систему.
В последнее время для выяснения роли базальных ганглиев в осуществлении движений успешно используют данные микроэлектродных исследований. Эксперименты на обезьянах показали наличие корреляции между разрядами нейронов полосатою тела и медленными, направленными из стороны в сторону червеобразными движениями лапы. Как правило, разряд нейрона предшествует началу медленного движения, а при быстрых "баллистических" движениях он отсутствует. Эти факты позволяют заключить, что нейроны полосатого тела участвуют в генерации медленных движений, подвергающихся коррекции со стороны сенсорной обратной связи.
По современным представлениям, базальныё ганглии являются одним из уровней построенной по иерархическому принципу системы регуляции движений.
Получая информацию от ассоциативных зон коры, базальныё ганглии участвуют в создании программы целенаправленных движений с учетом доминирующей мотивации. Далее соответствующая информация от базальных ганглиев поступает в передний таламус, где она интегрируется с информацией, приходящей от мозжечка. Из таламических ядер импульсация достигает двигательной коры, которая отвечает за реализацию программы целенаправленного движения через посредство нижележащих стволовых и спинальных двигательных центров. Так в общих чертах можно представить себе место базальных ганглиев в целостной системе двигательных центров мозга. Однако детальное исследование координирующей функции стриопаллидарной системы еще ждет своего разрешения.
При ревматическом поражении базальных ганглиев наблюдаются гиперкинезы — непроизвольные стереотипные повторяющиеся движения с разным ритмом и амплитудой . Эти движения особенно выражены у конечностей и проявляются в покое. Нарушается также регуляция тонуса скелетных мышц.
Важное значение в понимании роли функционирования базальных ганглиев имело выяснение их афферентных и эфферентных связей.
Афферентные волокна идут главным образом к полосатому телу и передают импульсы от трехисточников: всех отделов коры мозга, таламуса и черной субстанции . После обработки в полосатом теле эти импульсы идут к бледному шару и черной субстанции. Между нею и полосатым телом имеются двусторонние связи.
Черная субстанция влияет на нейроны базальных ганглиев с помощью медиатора дофамина. При дегенерации дофаминергических нейронов, нарушении синтеза и выделения этого медиатора нарушаются и функции системы базальных ганглиев. Появляются симптомы заболевания, называемого паркинсонизмом (дрожательный паралич). Среди симптомов паркинсонизма выделяют гипокинезию — резкое затруднение двигательных реакций, особенно их начала. Для таких больных характерна осторожная походка мелкими шажками. Появляется тремор покоя особенно выраженный на кистях рук. Этот тремор исчезает при осуществлении произвольного движения (в отличие от тремора, вызванного повреждением мозжечка, проявляющегося во время движений)
52. Функции коры больших полушарий. Сенсорные, моторные и ассоциативные зоны коры. Современные представления о локализации функций коры. Функциональная асимметрия коры у человека. Ее роль в реализации высших психический функций.
Кора головного мозга функционально состоит из трех зон: сенсорная зона, моторная зона и ассоциативная зона. У человека ассоциативная зона занимает около 75% коры головного мозга. Функция ассоциативной зоны - связывать между собой активность сенсорных и моторных зон. Ассоциативная зона, предполагается, получает и перерабатывает информацию из сенсорной зоны и инициирует целенаправленное осмысленное поведение.
Экспериментальными исследованиями установлено наличие в коре мозга трех зон, связанных со специфическими функциями и участками тела- двигательных, сенсорных и ассоциативных.Взаимосвязь между зонами позволяет координировать произвольные и непроизвольные формы деятельности, а также психические функции личности.
Павлов рассматривал кору большого мозга как совокупность корковых концов анализаторов. Корковый конец анализатора имеет не строго ограниченные границы, а состоит из ядерной и рассеянной частей. Ядро представляет собой точную проекцию в коре периферических рецепторов данной области и является необходимым для осуществления общего анализа и синтеза. Рассеянные элементы находятся по периферии ядра или могут быть разбросаны далеко от него. В них осуществляются более простые анализ и синтез.
Сенсорные зоны.
Сенсорные зоны занимают участки мозга, связанные с определенными видами чувствительности. В эти зоны поступает сенсорная информация.
Первичные сенсорные зоны- это области сенсорной коры, раздражение или разрушение которых вызывает четкие, постоянные изменения чувствительности организма (ядра анализаторов по Павлову).
Вокруг первичных зон находятся менее локализованные вторичные сенсорные зоны, нейроны которых отвечают на действие нескольких раздражителей.
Рассмотрим основные первичные зоны коры.
1) зона кожно-мышечной чувствительности (соматосенсорная зона)– теменная кора, пост(задне)-центральная извилина, поля 1,2,3,5,7 – к этой зоне приходят проприоцептивные импульсы от скелетных мышц, а также импулься от тактильных, температурных и других рецепторов кожи. Самую большую площадь зоны занимает область кисти, голосового аппарата, головы. Наименьшую площадь занимают представительства туловища, нижних конечностей. При повреждении зоны нарушается кожно-мышечная чувствительность