ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 05.12.2023
Просмотров: 3340
Скачиваний: 2
СОДЕРЖАНИЕ
Наиболее широко в ЦНС распространены медиаторы - амины:
Другие производные аминокислот - ГАМК, глицин, глютамин и др.
Название рецептора определено медиатором, с которым он взаимодействует:
Взаимодействие гормонов и парагормонов с клетками-мишенями
Понятие высших психических функций (Выготский)
Физиология газообмена в легких
Гуморальная, рефлекторная, нервная регуляция деятельности сердца
1.Общие свойства возбудимых тканей. Процесс возбуждения. Особенности местного и распространяющегося
Механизм формирования ПС связан с:
4. Современные представления о процессе возбуждения. Потенциал действия, его фазы.
8. Механизмы проведения возбуждения по нервным волокнам. Факторы, влияющие на скорость проведения
Механизмы проведения возбуждения по безмиелиновых нервным волокнам такой.
Закономерности проведения возбуждения через нервно-мышечный синапс:
В зависимости от частоты стимуляции выделяют следующие виды мышечного сокращения:
Тетанические сокращения отличается от одиночного следующими параметрами:
12. Функциональная характеристика гладких мышц.
13. Сила и работа мышц. Утомление и его особенности в целостном организме.
14. Нейрон как структурная и функциональная единица ЦНС. Его свойства и функции.
17-18. Возбуждение в ЦНС. Механизмы и закономерности передачи возбуждения в центральных
Особенности передачи возбуждения через центральные аксо-соматические химические синапсы.
Постсинаптическое гиперполяризацийне торможения.
Пресинаптическое деполяризации торможения.
Особенности передачи возбуждения в ЦНС:
Рефлекторная дуга имеет следующие звенья:
24. Рефлекс как элементарный акт нервной регуляции. Строение рефлекторной дуги
По расположению рецепторы подразделяют на:
По виду адекватного раздражителя, воспринимают рецепторы, их подразделяют на:
Физиологические механизмы кодирования информации в рецепторах.
26. Механизм кодирования информации в рецепторах. Адаптация рецепторов.
27. Общие принципы координационной деятельности ЦНС.
28. Суммация возбуждения, торможение нейронами ЦНС. Виды суммации и их значение
В зависимости от локализации рецепторного звена и эффекторного органа рефлексы делят на висцеро-
34. Сегментарные и надсегментарные центры вегетативной нервной системы
35. Гуморальная регуляция, её отличие от нервной. Факторы гуморальной регуляции.
Факторы гуморальной регуляции:
Механизм действия на клетки жирорастворимых гормонов:
Механизм действия жирорастворимых гормонов определяет следующие их особенности:
При воздействии на клетки-мишени водорастворимых гормонов образуются внутриклеточные посредники:
Классификация условных и безусловных рефлексов
- постоянство внутренней среды организма;
Современные представления о путях замыкания временных связей:
Эмоции выполнѐят две функции : сигнальную и регуляторную.
Эмоции делят на низшие и высшие.
Структурное обеспечение эмоций. Эмоциогенные структуры мозга.
Две сигнальные системы действительности
Типы высшей нервной деятельности
Общая характеристика восприятия
Безазотистые органические компоненты крови
Основные физико-химические константы крови:
Противосвертывающая система крови.
В норме гемоглобин содержится в виде нескольких соединений:
Методы исследования вентиляции легких:
Кислородная емкость крови, анализ кривой диссоциации:
Анализ кривой диссоциации НbО2:
^ Рефлекторная регуляция дыхания
второго порядка. Центр второго порядка может вырабатывать 40 - 60 импульсов в минуту.
^ Внутрисердечные механизмы регуляции.
Капиллярный кровоток и его особенности. Микроциркуляция и ее роль в механизме обмена жидкости и
Рефлекторная регуляция сердечно-сосудистой системы в зависимости от изменения положения тела в
Механизмы клубочковой фильтрации. Фильтрационное давление и факторы его определяющего. Состав
Механизм поддержания почками постоянства внутренней среды организма : рН, осмотического давления,
97. Функциональная система питания и пищеварения, ее основные звенья. Сенсорное насыщение. Функции
Пищеварение в полости рта. Состав и физиологическая роль слюны. Слюноотделение, его регуляция
101. Физиологическая роль печени, участие желчи в пищеварении. Факторы стимулирующие секрецию желчи,
106. Физиология щитовидной и околощитовидной желез
107. Физиология надпочечников. Роль гормонов коры и мозгового вещества в регуляции функции организма
Физиологическая характеристика обонятельной сенсорной системы. Механизмы восприятия запахов
Скорость доставки кислорода в нормальных условиях значительно превышает его потребление, в результате чего лишь малая доля доступного кислорода извлекается из капиллярной крови в обычном состоянии (в покое КУО2 = 22-32%). Это позволяет тканям приспосабливаться к снижению доставки кислорода увеличением его утилизации. КУО2 при тяжёлой мышечной работе способен повышаться до 60-80%.
70. Транспорт кислорода кровью. Кривая диссоциации оксигемоглобина, ее характеристика. Кислородная емкость крови.
Кислород, поступающий в кровь, сначала растворяется в плазме крови. При РАО, 100 мм рт. ст. в 100 мл плазмы растворяется 0,3 мл 02.
Кислород, растворился в плазме крови, по градиенту концентрации проходит через мембрану эритроцита и образует оксигемоглобин (НЬ02). При этом валентность железа не изменяется. Оксигемоглобин - неустойчивая соединение и легко разлагается. Прямая реакция называется оксигенацией, а обратный процесс - дезоксигенациею гемоглобина. При сочетании 02 с гемоглобином Fe2 + остается двухвалентным.
Каждая молекула НЬ может присоединить 4 молекулы 02, в пересчете на 1 г НЬ означает 1,34 мл 02. Зная количество гемоглобина в крови, можно определить кислородную емкость крови (КЕК): КЕК = НЬ-1, 34. Если в 100 мл крови содержится 15 г НЬ, то 15-1,34 = 20 мл 02 в 100 мл крови.
Учитывая, что 100 мл крови содержат только 0,3 мл растворенного 02, можно представить, что основной объем кислорода транспортируется в состоянии химической связи с гемоглобином. Но, несмотря на относительно низкую растворимость, количество растворенного в крови 02 можно увеличить искусственно. Растворимость газа в жидкости зависит от температуры, состава жидкости, давления газа и его природы. Поскольку состав крови, ее температура в организме почти всегда постоянны, количество растворенного газа можно вычислить по формуле:
Q = g • V • РаО2: Ратм, где Q-количество растворенного в жидкости газа; g - его адсорбционный коэффициент при t = 37 ° C (для 02 он составляет 0,023); V - объем крови, Ратм - атмосферное давление.
Когда увеличивается давление газа над жидкостью, количество растворенного газа увеличивается. Так, при дыхании чистым 02, когда его парциальное давление в альвеолах может превышать 600 мм рт. ст., в 100 мл крови растворяется уже около 2 мл кислорода. Но если человек находится в условиях с повышенным давлением кислорода (в барокамере), то количество растворенного в крови кислорода будет расти пропорционально давлению (гипербарическая оксигенация). Например, при парциальном давлении 3 атм, когда РАО, увеличивается до 2280 мм рт. ст. (304 кПа), в 100 мл крови может раствориться около 5-6 мл 02. Этого количества кислорода достаточно для того, чтобы ткани не испытывали кислородного даже при отсутствии связанного с гемоглобином 02. Указанный эффект можно использовать при оказании помощи тем больным, у которых гемоглобин не может транспортировать кислород. Например, дыхание чистым кислородом рекомендована при отравлении угарным газом, когда образуется стойкое соединение карбоксигемоглобин (диссоциирует в 1000 раз медленнее, чем оксигемоглобин).
Растворимость газов уменьшается при повышении температуры, но в условиях организма это большой роли не играет. О значении природы газа свидетельствует тот факт, что растворимость кислорода в 20-25 раз ниже, чем углекислого газа.
Зависимость насыщения гемоглобина O2 от парциального напряжения O2 может быть представлена графически в виде кривой диссоциации оксигемоглобина (рис. 711291326).
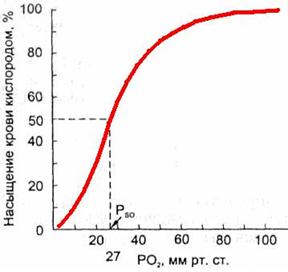
Рис. 711291326. Кривая диссоциации оксигемоглобина.
Кривая имеет сигмовидную форму, при этом нижняя часть кривой (РO2 < 60 мм рт.ст.) имеет крутой наклон, а верхняя часть (РO2 > 60 мм рт.ст.) относительно пологая.
Положение кривой диссоциации оксигемоглобина зависит от сродства гемоглобина с кислородом. При снижении сродства гемоглобина к O2, т.е. облегчении перехода O2 в ткани, кривая сдвигается вправо.
Повышение сродства гемоглобина к O2 означает меньшее высвобождение кислорода в тканях, при этом кривая диссоциации сдвигается влево.
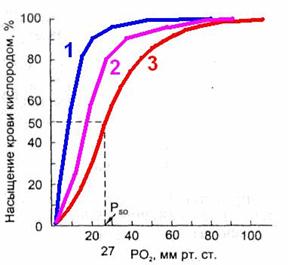
Важным показателем, отражающем сдвиги кривой диссоциации оксигемоглобина, является параметр Р50, т.е. такое РO2 , при котором гемоглобин насыщен кислородом на 50 % (рис. 7112913267).
В нормальных условиях у человека (при t 37 °С, рН 7,40 и РСO2= 40 мм рт.ст.) Р50 = 27 мм рт.ст.
При сдвиге кривой диссоциации вправо Р50 увеличивается, а при сдвиге влево — снижается.
На сродство гемоглобина к O2 оказывают влияние большое количество метаболических факторов, к числу которых относятся рН, РСO2, температура, концентрация в эритроцитах 2,3-дифосфоглицерата (2,3-ДФГ) (рис. 711291336).
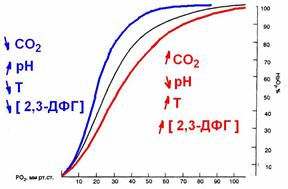
Рис. 711291336. Сдвиги кривой диссоциации оксигемоглобина.
Снижение рН, повышение РСО2 и температуры снижают сродство гемоглобина к О2 и смещению кривой вправо. Такие метаболические условия создаются в работающих мышцах, и такой сдвиг кривой является физиологически выгодным, так как повышенное высвобождение О2 необходимо для
Кислородная емкость крови — максимальное количество кислорода, которое может быть связано кровью. В среднем 1 г гемоглобина связывает около 1,35 см3 кислорода. Поэтому кислородная емкость крови зависит не от функции внешнего дыхания, а от содержания гемоглобина. Содержание кислорода в крови также зависит не только от эффективности вентиляции, диффузии и газообмена в легких, но и от содержания гемоглобина в крови. Весьма чувствительными показателями считаются парциальное давление (напряжение) кислорода и углекислоты. Определение парциального давления С02 может быть проведено вместе с измерением рН крови по методу Аструпа. Что же касается измерения парциального давления кислорода крови, то методика его сложна и вследствие этого не получила распространения в клинической практике. Наибольшее практическое значение вполне оправданно отводится определению степени насыщения крови кислородом, методика которого в настоящее время значительно усовершенствована, и это исследование получило широкое распространение в клинике торакальной хирургии. Методы определения степени насыщения крови кислородом разделяются на газометрические (манометрические) и оксигемометрические (спектрофотометрические). К первым относятся методы Ван-Слайка и Баркрофта. Газометрический способ Ван-Слайка основывается на принципах И. М. Сеченова — извлечение газов из крови в вакууме — и Холдейна — вытеснение газов химическими реактивами. Исследование газов крови на аппарате Ван-Слайка получило широкое распространение в клинической физиологии. К числу достоинств этого метода относится высокая точность результатов и возможность определения содержания кислорода и углекислоты. Однако длительность и трудоемкость исследования, необходимость относительно большого количества крови (1 мл), для чего требуется пункция артерии, ограничивают его применение в практической работе легочного хирурга, тем более, что определение наиболее важного показателя — степени насыщения крови кислородом — в настоящее время обычно производится с помощью оксигемометрии — метода, значительно более доступного и мало уступающего в точности
71. Транспорт углекислоты кровью, количество и формы ее содержания в крови. Роль эритроцитов в связывании и транспорте СО2.
В венозной крови содержится около 580 мл / л С02. В крови он содержится в трех формах: связанный в виде угольной кислоты и ее солей, связанный с гемоглобином и в растворенном виде.
С02 образуется в тканях при окислительных процессах. В большинстве тканей Рсо2 составляет 50-60 мм рт. ст. (6,7-8 кПа). В крови, поступающей в артериальное конец капилляров, РаCO2 составляет около 40 мм рт. ст. (5,3 кПа). Наличие градиента заставляет С02 диффундировать из тканевой жидкости до капилляров. Чем активнее в тканях осуществляются процессы окисления, тем больше создается СОТ и тем больше Ртк.со2. Интенсивность окисления в различных тканях различна. В венозной крови, оттекающей от ткани, Pvco приближается к 50 мм рт. ст. (6,7 кПа). А в крови, оттекающей от почек, Pvco2 составляет около 43 мм рт. ст. Поэтому в смешанной венозной крови, поступающей в правого предсердия, в состоянии покоя Pvco2 равна 46 мм рт. ст. (6,1 кПа).
С02 растворяется в жидкостях активнее, чем 02. При РCO2 равный 40 мм рт. ст. (5,3 кПа), в 100 мл крови растворено 2,4-2,5 мл СОГ, что составляет примерно 5% от общего количества газа, который транспортируется кровью. Кровь, проходящая через легкие, отдает далеко не весь С02. Большая часть его остается в артериальной крови, поскольку соединения, которые образуются на основе С02, участвуют в поддержании кислотно-основного равновесия крови - одного из параметров гомеостаза.
Химически связанный С02 находится в крови в одной из трех форм:
1) угольная кислота (Н2С03):
2) бикарбонатный ион (НСОИ)
3) карбогемоглобин (ННЬС02).
В форме угольной кислоты переносится только 7% СОГ, бикарбонатных ионов - 70%, карбогемоглобин - 23%.
С02, который проникает в кровь, сначала подвергается гидратации с образованием угольной кислоты: С02 + Н20 Н2СОз.
Эта реакция в плазме крови происходит медленно. В эритроците, куда С02 проникает по градиенту концентрации, благодаря специальному ферменту - карбоангидразы - этот процесс ускоряется примерно в 10 000 раз. Поэтому эта реакция происходит в основном в эритроцитах. Создаваемая здесь угольная кислота быстро диссоциирует на Н + и НСО3-, чему способствует постоянное образование угольной кислоты: Н2С03 Н + + НСО3-.
При накоплении НСО3-в эритроцитах создается его градиент с плазмой. Возможность выхода НСО3-в плазму определяется
условий: выход НСО3-должен сопровождаться одновременным выходом катиона или поступлением другого аниона. Мембрана эритроцита хорошо пропускает отрицательные, но плохо - положительные
ионы. Чаще образования и выход НСО3-из эритроцитов сопровождается поступлением в клетку СИ "". Это перемещение называют хлоридным сдвигом.
В плазме крови НСО3-"взаимодействуя с катионами, создает соли угольной кислоты. В виде солей угольной кислоты транспортируется около 510 мл / л С02.
Кроме того, СОТ может связываться с белками: частично - с белками плазмы, но главным образом - с гемоглобином эритроцитов. При этом сог взаимодействует с белковой частью гемоглобина - глобина. Гем же остается свободным и сохраняет способность гемоглобина находиться одновременно в связи как с С02, так и 02. Таким образом, одна молекула НЬ может транспортировать оба газа.
В крови альвеолярных капилляров все процессы осуществляются в противоположном направлении. Главная из химических реакций - дегидратация - происходит в эритроцитах при участии той же карбоангидразы: Н + + НСО3 Н2С03 Н20 + С02.
Направление реакции определяется непрерывным выходом С02 с эритроцита в плазму, а из плазмы в альвеолы. В легких в связи с постоянным его выделением происходит реакция диссоциации карбогемоглобин:
ННЬС02 +02 ННЬ02 + С02-> НЬ02 + Н + + С02.
Взаимосвязь транспорта кислорода и диоксида углерода. Выше указывалось, что форма кривой диссоциации оксигемоглобина влияет на содержание С02 в крови. Эта зависимость связана с тем, что дезоксигемоглобином является слабой кислотой, чем оксигемоглобин, и может присоединять более Н + Вследствие этого при уменьшении содержания оксигемоглобина повышается степень диссоциации Н2СОз, а следовательно, увеличивается транспорт С02 кровью. Эта зависимость называется эффектом Холдейна.
Взаимосвязь обмена двуокиси углерода и кислорода ярко обнаруживается в тканях и легких. К тканям поступает оксигенированный кровь. Здесь под влиянием С02 усиливается диссоциация гемоглобина. Поэтому поступление кислорода в ткани способствует ускорению поглощения С02 кровью.
В легких происходят обратные процессы. Поступление 02 снижает сродство крови к С02 и облегчает диффузию С02 в альвеолы. Это, в свою очередь, активизирует ассоциации гемоглобина с кислородом.
В то время как транспорт кислорода из легких к тканям почти полностью зависит от гемоглобина в эритроцитах, транспорт двуокиси (диоксида) углерода в обратном направлении немного сложнее. Двуокись углерода, в отличии от кислорода, растворима в плазме крови, так что большое количество СО2 переносится просто в растворенном виде. Остаток транспортируется эритроцитами. В тканях СО