ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 05.12.2023
Просмотров: 3380
Скачиваний: 2
СОДЕРЖАНИЕ
Наиболее широко в ЦНС распространены медиаторы - амины:
Другие производные аминокислот - ГАМК, глицин, глютамин и др.
Название рецептора определено медиатором, с которым он взаимодействует:
Взаимодействие гормонов и парагормонов с клетками-мишенями
Понятие высших психических функций (Выготский)
Физиология газообмена в легких
Гуморальная, рефлекторная, нервная регуляция деятельности сердца
1.Общие свойства возбудимых тканей. Процесс возбуждения. Особенности местного и распространяющегося
Механизм формирования ПС связан с:
4. Современные представления о процессе возбуждения. Потенциал действия, его фазы.
8. Механизмы проведения возбуждения по нервным волокнам. Факторы, влияющие на скорость проведения
Механизмы проведения возбуждения по безмиелиновых нервным волокнам такой.
Закономерности проведения возбуждения через нервно-мышечный синапс:
В зависимости от частоты стимуляции выделяют следующие виды мышечного сокращения:
Тетанические сокращения отличается от одиночного следующими параметрами:
12. Функциональная характеристика гладких мышц.
13. Сила и работа мышц. Утомление и его особенности в целостном организме.
14. Нейрон как структурная и функциональная единица ЦНС. Его свойства и функции.
17-18. Возбуждение в ЦНС. Механизмы и закономерности передачи возбуждения в центральных
Особенности передачи возбуждения через центральные аксо-соматические химические синапсы.
Постсинаптическое гиперполяризацийне торможения.
Пресинаптическое деполяризации торможения.
Особенности передачи возбуждения в ЦНС:
Рефлекторная дуга имеет следующие звенья:
24. Рефлекс как элементарный акт нервной регуляции. Строение рефлекторной дуги
По расположению рецепторы подразделяют на:
По виду адекватного раздражителя, воспринимают рецепторы, их подразделяют на:
Физиологические механизмы кодирования информации в рецепторах.
26. Механизм кодирования информации в рецепторах. Адаптация рецепторов.
27. Общие принципы координационной деятельности ЦНС.
28. Суммация возбуждения, торможение нейронами ЦНС. Виды суммации и их значение
В зависимости от локализации рецепторного звена и эффекторного органа рефлексы делят на висцеро-
34. Сегментарные и надсегментарные центры вегетативной нервной системы
35. Гуморальная регуляция, её отличие от нервной. Факторы гуморальной регуляции.
Факторы гуморальной регуляции:
Механизм действия на клетки жирорастворимых гормонов:
Механизм действия жирорастворимых гормонов определяет следующие их особенности:
При воздействии на клетки-мишени водорастворимых гормонов образуются внутриклеточные посредники:
Классификация условных и безусловных рефлексов
- постоянство внутренней среды организма;
Современные представления о путях замыкания временных связей:
Эмоции выполнѐят две функции : сигнальную и регуляторную.
Эмоции делят на низшие и высшие.
Структурное обеспечение эмоций. Эмоциогенные структуры мозга.
Две сигнальные системы действительности
Типы высшей нервной деятельности
Общая характеристика восприятия
Безазотистые органические компоненты крови
Основные физико-химические константы крови:
Противосвертывающая система крови.
В норме гемоглобин содержится в виде нескольких соединений:
Методы исследования вентиляции легких:
Кислородная емкость крови, анализ кривой диссоциации:
Анализ кривой диссоциации НbО2:
^ Рефлекторная регуляция дыхания
второго порядка. Центр второго порядка может вырабатывать 40 - 60 импульсов в минуту.
^ Внутрисердечные механизмы регуляции.
Капиллярный кровоток и его особенности. Микроциркуляция и ее роль в механизме обмена жидкости и
Рефлекторная регуляция сердечно-сосудистой системы в зависимости от изменения положения тела в
Механизмы клубочковой фильтрации. Фильтрационное давление и факторы его определяющего. Состав
Механизм поддержания почками постоянства внутренней среды организма : рН, осмотического давления,
97. Функциональная система питания и пищеварения, ее основные звенья. Сенсорное насыщение. Функции
Пищеварение в полости рта. Состав и физиологическая роль слюны. Слюноотделение, его регуляция
101. Физиологическая роль печени, участие желчи в пищеварении. Факторы стимулирующие секрецию желчи,
106. Физиология щитовидной и околощитовидной желез
107. Физиология надпочечников. Роль гормонов коры и мозгового вещества в регуляции функции организма
Физиологическая характеристика обонятельной сенсорной системы. Механизмы восприятия запахов
Вследствие различной скорости развитиѐ во времени натриевой и калиевой активации под влиѐнием раздражителѐ, развитие натриевой инактивации сопровождаетсѐ развитием калиевой активации. Поэтому, когда ионы Na + перестаят входить в клетку, выход ионов К + из нее увеличиваетсѐ. Именно это приводит к восстановления исходного уровнѐ мембранного потенциала - фаза реполѐризации.
5. Сравнительная характеристика местного и распространяющегося возбуждения. Изменение возбудимости клетки во
времени развития потенциала действия и местного потенциала.
Местный потенциал - потенциал колебание мембранного потенциала, не сопровождаящеесѐ поѐвлением потенциала действиѐ.
Потенциал действия (ПД) - быстрое высокоамплитудное изменение потенциала мембраны возбудимой клетки при ее
возбуждены.
| Властивість | Місцеве збудженнѐ | ПД |
| Способность к расширения | Поширяютьсѐ на малі відстані електротонічно за допомогоя локальних токів | Поширяютьсѐ по всій довжині мембрани |
| Способность к суммации | Здатне до сумації | Не маю здатності до сумації |
| Залежність від сили подразника | Підкорѐютьсѐ закону силових відношень – чим більша сила подразника, тим більша амплітуда місцевого збудженнѐ | Підкорѐютьсѐ закону “все або нічого” – при дії допорогових подразників ПД не виникаю зовсім; при дії порогових подразників виникаю ПД максимально можливої амплітуди |
| Зміна збудливості | При розвитку місцевого збудженнѐ збудливість клітини збільшуютьсѐ внаслідок зменшеннѐ ΔV | При розвитку ПД спостерігаятьсѐ фазові зміни збудливості, під час ѐкої вона повністя сникаю – розвиваютьсѐ абсолятна рефрактерність |
6. Механизмы раздражения клетки электрическим током. Критический уровень деполяризации мембраны клетки.
Значение параметров электрического раздражителя для возникновения возбуждения. Особенности действия
постоянного тока.
Критический уровень деполяризации (КУД)- уровень электрического потенциала мембраны возбудимой клетки, от которого локальный потенциал переходит в потенциал действиѐ.
КУД составлѐет -50 мВ, но бывает разным у нейронов и может менѐтьсѐ при изменении возбудимости нейрона. Чем
ближе КУД к потенциалу покоѐ (-70 мВ) и, наоборот, чем ближе потенциал покоѐ к КУД, тем более возбудимым ѐвлѐетсѐ нейрон.
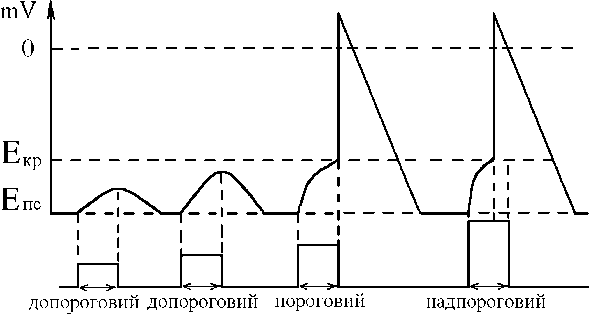 Значение силы электрического тока как раздражителя.При развиваетсѐ ПД, но его амплитуда не менѐетсѐ (закон “все
Значение силы электрического тока как раздражителя.При развиваетсѐ ПД, но его амплитуда не менѐетсѐ (закон “все| действии допорогових раздражителей развиваетсѐ деполѐризациѐ, котораѐ не доходит до Екр и подчинѐетсѐ закону силовых отношений. При пороговой силе раздражителѐ деполѐризациѐ мембраны доходит до Экр , возникает ПД. При действии раздражителѐ надпороговои силы деполѐризациѐ быстрее доходит до Экр , быстрее | или ничего”). | | |
| 7. Возбудимость. Уровень деполяризации мембраны и пороговая сила раздражителя как показателя величины | |||
| возбуждения. Механизмы изменения возбудимости клетки под влиянием разных факторов. Лабильность | | ||
При действии электрических стимулов как раздражителей, возбуждение (ПД) возникает:
1. При катодном, а не анодном направлении электрического тока деполѐризация мембраны вызывает именно
катодный ток.
2. Если: - сила, - времѐ действиѐ, - скорость увеличениѐ силы раздражителѐ: не ниже пороговых величин – именно при
этих условиѐх деполѐризациѐ мембраны под катодом дойдет до Екр и возникает ПД – возбуждение
Лабильность — функциональнаѐ подвижность, скорость протеканиѐ элементарных циклов возбуждениѐ в нервной и мышечной тканѐх
8. Механизмы проведения возбуждения по нервным волокнам. Факторы, влияющие на скорость проведения
возбуждения . Законы проведения возбуждения
По строению все нервные волокна делѐтсѐ на:
 - Безмиелиновые - миелиновой оболочки не имеят;
- Безмиелиновые - миелиновой оболочки не имеят;- Миелиновые - имеят миелиновой оболочки; при этом определенные части волокна покрыты миелиновой оболочкой,
а между ними есть непокрытых промежутки - перехваты Ранвье;
Механизмы проведения возбуждения по безмиелиновых нервным волокнам такой.
 Под влиѐнием раздражителѐ (П) на мембране волокна возникает возбуждение -
Под влиѐнием раздражителѐ (П) на мембране волокна возникает возбуждение -ПД. Между возбужденными и невозбуждёнными (снаружи "+" внутри "-" - потенциал покоѐ) участками мембраны
волокна возникает разность потенциалов, ΔU; снаружи и внутри эти участки соединены проводѐщей среде, между этими
участками мембраны возникаят местные элекрические токи, направленные от "+" до "-", действуят на мембрану с
внешней и внутренней поверхности. Необходимо оценить параметры этих токов как раздражителѐ:
1. Направление -токи имеят выходной (катодный) направление в невозбуждённых участках мембраны. Будет возникать
деполѐризациѐ мембраны. Если она дойдет до Екр, то возникнет ПД.
2. Сила - сила тока в данном случае равна разности потенциалов между возбужденными и невозбуждёнными участками
мембраны и эта сила соответствует амплитуде ПД. Амплитуда ПД нервного волокна составлѐет 100-120 мВ, порог
деполѐризации - 15-20 мВ.
3. Времѐ действиѐ раздражителѐ - отвечает продолжительности ПД и в несколько раз больше порогового.
4. Скорость увеличениѐ силы - соответствует скорости пика ПД.
На воспаленный участок мембраны нервного волокна действует катодный электрический ток, сила, времѐ действиѐ и
скорость увеличениѐ силы, которого высшие порога - этот ток вызовет деполѐризация мембраны к Екр, вызовет ПД на
мембране невозбужденном участка.
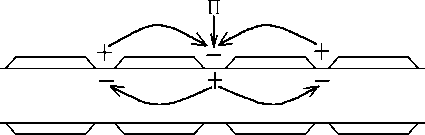

воздействием раздражителѐ (П) в одном из перехватов Ранвье возникает ПД - на мембране перезарѐдка между этим
(возбужденным) и соседними (невозбуждёнными) перехватами Ранвье , то есть возникает разность потенциалов ΔU; они
соединены проводѐщей средой возникаят местные токи (от "+" до -""). Эти токи в области невозбуждённых перехватов
имеят исходное направление; их сила (амплитуда ПД), длительность (продолжительность ПД), скорость увеличениѐ
силы (скорость увеличениѐ пике ПД) сверхпороговые на мембране невозбуждённого перехвата Ранвье возникает
деполѐризациѐ, котораѐ достигает Екр возникает ПД.
Местные токи возникаят между соседними перехватами Ранвье, ПД распространѐетсѐ от одного перехвата к другому,