ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 05.12.2023
Просмотров: 3416
Скачиваний: 2
СОДЕРЖАНИЕ
Наиболее широко в ЦНС распространены медиаторы - амины:
Другие производные аминокислот - ГАМК, глицин, глютамин и др.
Название рецептора определено медиатором, с которым он взаимодействует:
Взаимодействие гормонов и парагормонов с клетками-мишенями
Понятие высших психических функций (Выготский)
Физиология газообмена в легких
Гуморальная, рефлекторная, нервная регуляция деятельности сердца
1.Общие свойства возбудимых тканей. Процесс возбуждения. Особенности местного и распространяющегося
Механизм формирования ПС связан с:
4. Современные представления о процессе возбуждения. Потенциал действия, его фазы.
8. Механизмы проведения возбуждения по нервным волокнам. Факторы, влияющие на скорость проведения
Механизмы проведения возбуждения по безмиелиновых нервным волокнам такой.
Закономерности проведения возбуждения через нервно-мышечный синапс:
В зависимости от частоты стимуляции выделяют следующие виды мышечного сокращения:
Тетанические сокращения отличается от одиночного следующими параметрами:
12. Функциональная характеристика гладких мышц.
13. Сила и работа мышц. Утомление и его особенности в целостном организме.
14. Нейрон как структурная и функциональная единица ЦНС. Его свойства и функции.
17-18. Возбуждение в ЦНС. Механизмы и закономерности передачи возбуждения в центральных
Особенности передачи возбуждения через центральные аксо-соматические химические синапсы.
Постсинаптическое гиперполяризацийне торможения.
Пресинаптическое деполяризации торможения.
Особенности передачи возбуждения в ЦНС:
Рефлекторная дуга имеет следующие звенья:
24. Рефлекс как элементарный акт нервной регуляции. Строение рефлекторной дуги
По расположению рецепторы подразделяют на:
По виду адекватного раздражителя, воспринимают рецепторы, их подразделяют на:
Физиологические механизмы кодирования информации в рецепторах.
26. Механизм кодирования информации в рецепторах. Адаптация рецепторов.
27. Общие принципы координационной деятельности ЦНС.
28. Суммация возбуждения, торможение нейронами ЦНС. Виды суммации и их значение
В зависимости от локализации рецепторного звена и эффекторного органа рефлексы делят на висцеро-
34. Сегментарные и надсегментарные центры вегетативной нервной системы
35. Гуморальная регуляция, её отличие от нервной. Факторы гуморальной регуляции.
Факторы гуморальной регуляции:
Механизм действия на клетки жирорастворимых гормонов:
Механизм действия жирорастворимых гормонов определяет следующие их особенности:
При воздействии на клетки-мишени водорастворимых гормонов образуются внутриклеточные посредники:
Классификация условных и безусловных рефлексов
- постоянство внутренней среды организма;
Современные представления о путях замыкания временных связей:
Эмоции выполнѐят две функции : сигнальную и регуляторную.
Эмоции делят на низшие и высшие.
Структурное обеспечение эмоций. Эмоциогенные структуры мозга.
Две сигнальные системы действительности
Типы высшей нервной деятельности
Общая характеристика восприятия
Безазотистые органические компоненты крови
Основные физико-химические константы крови:
Противосвертывающая система крови.
В норме гемоглобин содержится в виде нескольких соединений:
Методы исследования вентиляции легких:
Кислородная емкость крови, анализ кривой диссоциации:
Анализ кривой диссоциации НbО2:
^ Рефлекторная регуляция дыхания
второго порядка. Центр второго порядка может вырабатывать 40 - 60 импульсов в минуту.
^ Внутрисердечные механизмы регуляции.
Капиллярный кровоток и его особенности. Микроциркуляция и ее роль в механизме обмена жидкости и
Рефлекторная регуляция сердечно-сосудистой системы в зависимости от изменения положения тела в
Механизмы клубочковой фильтрации. Фильтрационное давление и факторы его определяющего. Состав
Механизм поддержания почками постоянства внутренней среды организма : рН, осмотического давления,
97. Функциональная система питания и пищеварения, ее основные звенья. Сенсорное насыщение. Функции
Пищеварение в полости рта. Состав и физиологическая роль слюны. Слюноотделение, его регуляция
101. Физиологическая роль печени, участие желчи в пищеварении. Факторы стимулирующие секрецию желчи,
106. Физиология щитовидной и околощитовидной желез
107. Физиология надпочечников. Роль гормонов коры и мозгового вещества в регуляции функции организма
Физиологическая характеристика обонятельной сенсорной системы. Механизмы восприятия запахов
-
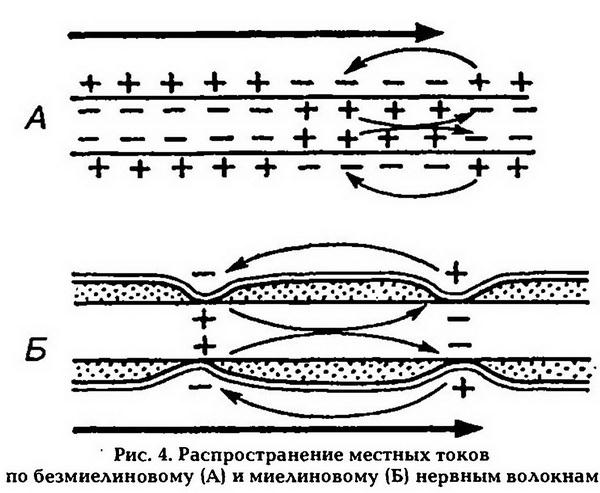
Да, так как ПД регистрируется в перехватах Ранвье, и распространяется по мембране волокна сальтаторно -
Отдельные участки волокна №1 покрыты миелином, имеющим свойства диэлектрика, поэтому в межперехватных участках ПД не регистрируется -
Волокна 1 и 2 имеют различные физические свойства, так как 1 волокно является миелиновым, а 2 безмиелиновым -
Участки волокна 1, где регистрируется ПД называется перехватом Ранвье, в этих участках отсутствует миелиновая оболочка, и большое количество натриевых каналов, активация которых приводит к деполяризации, а следовательно и к возникновению ПД -
Для любого нервного волокна общим механизмом проведения является двухстороннее проведение возбуждения, изолированное проведение возб-я по нервному волокну и анатомо-физиологическая целостность волокна -
В миелиновых волокнах (№1) возбуждение возникает пи нанесении раздр – я в перехватах Ранвье и распространаяется по мембране сальтаторно, при этом может охватывать и несколько перехватов, что является энергитически более выгодным и увеличивает скорость распространения возб-я. В безмиелиновых волокнах (№2) на мембране волокна между возб. И невозб. Участками возникает местный ток, который является раздражителем для соседнего невозб. Участка, но скорость передачи импульса резко снижается. -
32 При определении скорости проведения возб-я в двух нервных волокнах было установлено, что скор проведения в вол 1 составляет 100м/с, в вол 2 – 10 м/с.
-
Волокно 1 – миелиновое типа А наибольшего диаметра, скорость 120-70 м/с, длительность ПД минимальная
Волокно 2 – миелиновое типа В меньшего диаметра, скорость 3-18 м/с, ПД более длителен -
Cкорость проведения возбуждения в волокнах 1 и 2 зависит от диаметра волокна -
Скорость проведения возб-я в волокнах 1 и 2 прямо пропорциональна длине межперехватных участков, а их длина тем больше чем больше диаметр волокна.
33 При определении структурных и функциональных показателей двух афферентных волокон 1 и 2 было установлено, что диаметр волокна 1 - 20 мкм, после пика его пд, развивается гиперполяризация ( 50 мс), диаметр вол 2 - 2 мкм, гиперполяризация – 500 мс
-
Миелиновые и безмиелиновые -
Миелиновые волокна имеют миелиновую оболочку, а безмиелиновые только шванновскую оболочку. Миелиновые – диаметр до 25 мкм, скорость 120 м/с, продолжит ПД 0,4-0,5 мсек. Безмиелиновые – до 1,3 мкм, до 2 м/с, 2мсек Безмиелиновые (безмякотные) волокна покрыты только леммоцитами (шванновскими клетками). Между ними и осевым цилиндром (аксоном нейрона) имеется щель с межклеточной жидкостью, поэтому, клеточная мембрана остается неизолированной. Импульс распространяется по волокну со скоростью всего 1-3 м/сек.Миелиновые волокна покрыты спиральными слоями шванновских клеток с прослойкой миелина - жироподобного вещества с высоким удельным сопротивлением. Миелиновая оболочка через промежутки равной длины прерывается, оставляя оголенными участки осевого цилиндра длиной » 1 мкм -
Основные типы нервных волокон: Аα(первичные афференты мышечных веретен, двигательные волокна скелетным мышц) – 15 мкм, 70-120 м/с; Аβ ( кожные афференты прикосновения и давления)- 8мкм, 30-70 м/с; Аγ ( двигательные волокна мышечных веретен) 5мкм, 15-30 м/с; Аδ ( кожные афференты температуры и боли) < 3мкм, 12-30м/с. В(симпатические преганглионарные волокна) 3 мкм, 15 м/с. С( кожные афференты боли, симпатические постганглионарные волокна (немиелинизированные)) 1 мкм, 0,5-2 м/с -
Волокно 1 – миелиновое типа Аα, волокно 2 – миелиновые Аδ -
1- первичные афференты мышечных веретен, 2- кожные афференты температуры и боли -
Лабильность определяется длительностью рефрактерного периода(фаз абсолютной рефрактерности), чем длиннее рефрактерная фаза, тем меньше лабильность. -
Фаза абсолютной рефрактерности = фаза деполяризации(восходящая фаза ПД), -
У волокна 1 лабильность выше, так как короче рефрактерный период, у волокна 2 – наоборот
34 В эксперименте исследовали потенциалы двух нерв вол 1 и 2, идущих в сост-е смешанного нерва конечности. Группу вол 1 сост волокна, идущие от т.н. проприорецепторов, группу 2 – от болевых рецепторов кожи.
1,2,3. Группа 1 – волокна типа Аα, а группа 2 – Аδ, следовательно, волокна группы 1 имели наименьшую продолжительность ПД, а группы 2 – чуть более длительный ПД, что обусловлено различной длиной межперехватных участков(?)
-
Законы проведения вобз-я по нервам:
Закон анатомо-физиологической целостности волокна
Закон изолированного проведения возбуждения по нервному волокну
Закон двухстороннего проведения возбуждения
-
В двигательных волокнах скелетных мышц, которые так же относятся к типу Аα
35 Частота раздр-я нерва 500стимулов/с, возб – е походит через поврежд участок, в котором длительность развития пд в рез-те травмы увеличилась с 0.5 мс до 5 мс, след потенциалы не характерны. Сопоставить частоту пд, до поврежд участка и после него
-
Совпадает, т.к. чем выше частота раздражения – тем сильнее ответ(выше частота пд) -
В поврежденном участке возникают потенциалы меньшие по своей амплитуде, а значит и по частоте -
Наблюдается явление парабиоза-резкого снижения лабильности -
Лабильность - максимальное кол-во циклов возбуждения, которые может генерировать ткань в единицу времени. Отсюда следует, что неповрежденный участок обладает большей лабильностью(длительность ПД меньше, а значит и рефрактерный период короче) по сравнению с поврежденным участком. -
Так как парабиоз – нераспространяющееся возбуждение, то после поврежденного участка частота пд и раздражения совпадают (????)
36 При раздр нервной ткани был зарегистрирован пд, амплитуда к-рого по мере увеличения силы разд-я увеличивалась, а характер кривой не менялся.
-
Объектом был смешанный нерв. -
Если электрод расположен так, что контактирует с целым нервом, то при стимуляции нерва на некотором расстоянии от места отведения электрод сначала зарегистрирует потенциалы действия наиболее быстро проводящих волокон, а после этого–группы потенциалов других, более медленно проводящих волокон. Следовательно, потенциал действия такого нерва состоит из ответов целого спектра групп волокон с разными скоростями проведения.
37 При надпороговом раздр-ии двиг нерва были зарегистрированы его биопотенциалы ( ПД) и ПД иннерв-й скелетной мышцы. ПД мышцы не возникли при действии ионов Mg, явл антагонистами ионов Са
-
Возбуждение передается посредством синапса, его основные составляющие – пресинаптическая мембрана, синаптическая щель, постсинаптическая мембрана -
Mg не действует на первое звено передачи, так как не возник ПД на мышце. Значит Мg не действует на пресинаптическую мембрану нервного окончания. -
Не действует ни на одно звено -
Mg не оказывает действия на синапс, а Са действует на пресинаптическую мембрану -
Са играет ключевую роль в высвобождении квантов медиатора в синаптическую щель- -
Деполяризация пресинаптической мамбраны – открытие Са-каналов – высвобождение медиатора -
Высвобождается медиатор, который связывается с лиганд-зависимыми Na-каналами на постсинаптической мембране, открывая их – увеличение проницаемости для натрия – генерация пд на постсинаптической мембране
38 . При надпороговом раздр двиг нерва были зарег его биопотенциалы ( ПД) и ПД иннерв-й скелет мышцы. ПД мышцы не возн при действии магния, явл антагонистом Са. При прямом раздр мышцы она продолжала возбуждаться и сокращаться
-
Да -
Ионы магния не принимают участия в генерации пд -
Нет -
Так как мышца продолжала возбуждаться и сокращаться при ее прямом радражении -
-
39 В экспер регистрировали биопотенциалы нерв вол при его пороговом раздражении и потенц иннервируемого мыш волокна при прямом раздражении. Сила раздр в обоих случаях одинакова
40 В эксперименте на 2х мышцах ( а и б ) отдельное мыш волокно раздражали внутриклеточно надпороговым током. Было установлено, что на мышце а – регистр пд и других мыш волокон, на мышце б – пд только раздражаемого волокна
-
Между клетками волокон существует система межклеточных контактов(нексусы), через которые распространяется пд -
А – гладкая висцеральная мышца(унитарная), Б – скелетная -
В гладкой мышце нет концевых пластиной и отдельных нервных окончаний, по всей длине разветвлений адрен- и холинэргических нейронов имеются варикозные утолщения, содержацие гранулы медиатора. Поэтому по ходу следования нервного волокна могут возбуждаться многие гладкомышечные клетки, а клетки не имеющие контакта с варикозами возбуждаются посредством пд, распространяющегося на них через нексусы. -
В скелетной мышце нет связей между отдельными мышечными волокнами, поэтому пд не распространяется на соседние волокна
№
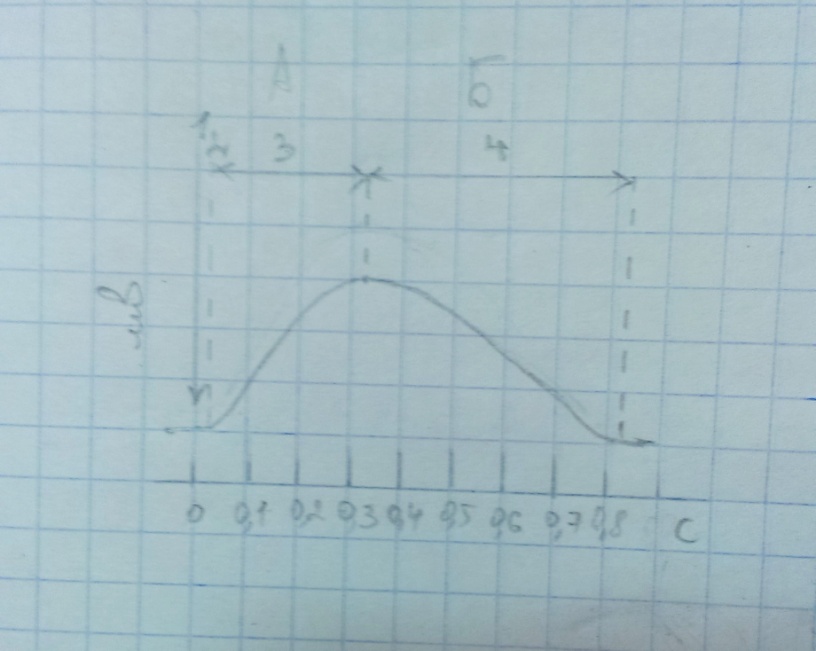 41. Задачей эксперимента являлось получение гладкого и зубчатого тетануса изолированной мышцы. Длительность латентного периода ее сокращения составляла 0.02с, периода укорочения – 0.30с, периода расслабления – 0.50с. Длительность фазы абсолютной рефрактерности данной мышцы – 0.35с, фазы относительной рефрактерности – 0.10с. Каковы результаты эксперимента?
41. Задачей эксперимента являлось получение гладкого и зубчатого тетануса изолированной мышцы. Длительность латентного периода ее сокращения составляла 0.02с, периода укорочения – 0.30с, периода расслабления – 0.50с. Длительность фазы абсолютной рефрактерности данной мышцы – 0.35с, фазы относительной рефрактерности – 0.10с. Каковы результаты эксперимента?
1. 1 – момент раздражения, 2 – латентный период, 3 – период укорочения, 4 – период расслабления.
2. Общее условие получения тетанического сокращения – действие на мышцу ритмических раздражений с такой частотой, что их эффекты суммируются, наступает сильное и длительное сокращение мышцы. ИЛИ наличие абсолютного рефрактерного периода (как сказано в методичке) (?)
3. При нанесении последующего раздражения во время укорочения возникает гладкий тетанус, а во время расслабления – зубчатый тетанус.
4. Минимальный промежуток времени между последовательными эффективными стимулами во время тетануса не может быть меньше рефрактерного периода. Следовательно, для данной мышцы возможно получение зубчатого тетануса (период расслабления > рефрактерный период), и невозможно получение гладкого (период укорочения < рефрактерный период).
5. Одинаковы.
6. При пороговой силе раздражителя ответ не будет наблюдаться ни во время фазы абсолютной рефрактерности, ни во время относительной. А при максимальной силе возможен будет ответ во время фазы относительной рефрактерности. Однако на частоту раздражения данной мышцы это не повлияет, так как останется: период расслабления > рефрактерный период и период укорочения < рефрактерный период.
№42. Сопоставьте динамическую работу, выполненную изолированной мышцей лягушки при однократном подъеме грузов в 10г, 30г, 410г, при условии, что абсолютная сила мышцы составляет 400г.
1. Динамическая работа – работа мышцы, при которой происходит перемещение груза и движение костей в суставах.
2
 . Учитывая, что абсолютная сила мышцы составляет 400г, а необходимо поднять груз 410г, то в данном случае выполненная работа будет равняться нулю. Поэтому при грузе 10г работа будет больше, чем при грузе 410г.
. Учитывая, что абсолютная сила мышцы составляет 400г, а необходимо поднять груз 410г, то в данном случае выполненная работа будет равняться нулю. Поэтому при грузе 10г работа будет больше, чем при грузе 410г.3. Работа при грузе 30г больше работы при грузе 10г. Высота подъема увеличилась.
4. Исходя из формулы для измерения динамической работы мышц (А = РН, где А – динамическая работа мышцы, кгм, Р – масса груза, кг, Н – высота подъема груза, м.), динамическая работа мышцы прямо пропорциональна массе груза и высоте подъема груза.
5. По мере увеличения груза работа сначала увеличивается, а затем постепенно уменьшается. При очень большом грузе, который мышца неспособна поднять, работа становится равной нулю. Наибольшую работу мышца совершает при некоторых средних нагрузках.
№43. На скелетной мышце были проведены 2 опыта (№1 и №2), в которых регистрировали ее потенциалы возбуждения (ПД). В опыте №1 на мышцах наносили прямое раздражение, в опыте №2 – непрямое. Частота раздражения в двух случаях была одинаковой и составляла 250 стимулов в секунду.
Будут ли наблюдаться в опытах №1 и №2 различия в частоте ПД, возникающие в мышце при условии, что лабильность нерва равна 500 имп/с, лабильность мышцы – 300 имп/с.
1. При прямом раздражении мышцы стимулами указанной частоты, примерная частота импульсов, возникающих в мышце –250 имп/с.
2. Поскольку частота прямых раздражений - 250 стимулов в секунду, а лабильность (способность ткани воспроизводить определенное число волн возбуждения в единицу времени в точном соответствии с ритмом наносимых раздражений) мышцы – 300 имп/с.
3. При частоте раздражения нерва 250 стимулов/с, в нерве будет возникать 250 имп/с, поскольку его лабильность - 500 имп/с.
4. При непрямом раздражении указанной частоты в мышце будет возникать около 100-150 имп/с.
5. В данном случае происходит раздражение двигательного нерва, возбуждение от которого передается на мышцу посредством синапса, лабильность которого 100-150 имп/с.
6. Лабильность структур, по которым проходит возбуждение при непрямом раздражении мышцы: нерв – 500имп/с, синапс – 100-150 имп/с, мышца - 300 имп/с.
№44. Животному было введено курареподобное вещество, действующее в области мышечно-нервного синапса скелетной мышцы. Как доказать точную локализацию действия веществ и его механизм?
1. После введения примененного вещества скелетная мышца не будет реагировать на непрямое надпороговое раздражение.
2. Чтобы доказать, что указанное вещество не действует на двигательный нерв, а именно в области синапса, нужно измерить ПД отведением с данного нерва.
3. Доказать, что данное вещество не действует на сократительный аппарат мышц можно прямым раздражением.
4. Проверить, не действует ли данное вещество в области пресинаптической мембраны необходимо, проверив наличие медиатора ацетилхолина в синаптической щели.
5. Проверить, не оказывает ли данное вещество антихолинэстеразное действие, можно измерив ПД постсинаптической мембраны, т.к. при действии ингибитора холинэстеразы ритмическое раздражение нерва вызывает суммацию потенциала концевой пластинки, что ведет к стойкой деполяризации постсинаптической мембраны. (?)
6. Обладает ли вещество деполяризующим действием нужно проверить, измерив ПД постсинаптической мембраны. (?)