ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 05.12.2023
Просмотров: 3418
Скачиваний: 2
СОДЕРЖАНИЕ
Наиболее широко в ЦНС распространены медиаторы - амины:
Другие производные аминокислот - ГАМК, глицин, глютамин и др.
Название рецептора определено медиатором, с которым он взаимодействует:
Взаимодействие гормонов и парагормонов с клетками-мишенями
Понятие высших психических функций (Выготский)
Физиология газообмена в легких
Гуморальная, рефлекторная, нервная регуляция деятельности сердца
1.Общие свойства возбудимых тканей. Процесс возбуждения. Особенности местного и распространяющегося
Механизм формирования ПС связан с:
4. Современные представления о процессе возбуждения. Потенциал действия, его фазы.
8. Механизмы проведения возбуждения по нервным волокнам. Факторы, влияющие на скорость проведения
Механизмы проведения возбуждения по безмиелиновых нервным волокнам такой.
Закономерности проведения возбуждения через нервно-мышечный синапс:
В зависимости от частоты стимуляции выделяют следующие виды мышечного сокращения:
Тетанические сокращения отличается от одиночного следующими параметрами:
12. Функциональная характеристика гладких мышц.
13. Сила и работа мышц. Утомление и его особенности в целостном организме.
14. Нейрон как структурная и функциональная единица ЦНС. Его свойства и функции.
17-18. Возбуждение в ЦНС. Механизмы и закономерности передачи возбуждения в центральных
Особенности передачи возбуждения через центральные аксо-соматические химические синапсы.
Постсинаптическое гиперполяризацийне торможения.
Пресинаптическое деполяризации торможения.
Особенности передачи возбуждения в ЦНС:
Рефлекторная дуга имеет следующие звенья:
24. Рефлекс как элементарный акт нервной регуляции. Строение рефлекторной дуги
По расположению рецепторы подразделяют на:
По виду адекватного раздражителя, воспринимают рецепторы, их подразделяют на:
Физиологические механизмы кодирования информации в рецепторах.
26. Механизм кодирования информации в рецепторах. Адаптация рецепторов.
27. Общие принципы координационной деятельности ЦНС.
28. Суммация возбуждения, торможение нейронами ЦНС. Виды суммации и их значение
В зависимости от локализации рецепторного звена и эффекторного органа рефлексы делят на висцеро-
34. Сегментарные и надсегментарные центры вегетативной нервной системы
35. Гуморальная регуляция, её отличие от нервной. Факторы гуморальной регуляции.
Факторы гуморальной регуляции:
Механизм действия на клетки жирорастворимых гормонов:
Механизм действия жирорастворимых гормонов определяет следующие их особенности:
При воздействии на клетки-мишени водорастворимых гормонов образуются внутриклеточные посредники:
Классификация условных и безусловных рефлексов
- постоянство внутренней среды организма;
Современные представления о путях замыкания временных связей:
Эмоции выполнѐят две функции : сигнальную и регуляторную.
Эмоции делят на низшие и высшие.
Структурное обеспечение эмоций. Эмоциогенные структуры мозга.
Две сигнальные системы действительности
Типы высшей нервной деятельности
Общая характеристика восприятия
Безазотистые органические компоненты крови
Основные физико-химические константы крови:
Противосвертывающая система крови.
В норме гемоглобин содержится в виде нескольких соединений:
Методы исследования вентиляции легких:
Кислородная емкость крови, анализ кривой диссоциации:
Анализ кривой диссоциации НbО2:
^ Рефлекторная регуляция дыхания
второго порядка. Центр второго порядка может вырабатывать 40 - 60 импульсов в минуту.
^ Внутрисердечные механизмы регуляции.
Капиллярный кровоток и его особенности. Микроциркуляция и ее роль в механизме обмена жидкости и
Рефлекторная регуляция сердечно-сосудистой системы в зависимости от изменения положения тела в
Механизмы клубочковой фильтрации. Фильтрационное давление и факторы его определяющего. Состав
Механизм поддержания почками постоянства внутренней среды организма : рН, осмотического давления,
97. Функциональная система питания и пищеварения, ее основные звенья. Сенсорное насыщение. Функции
Пищеварение в полости рта. Состав и физиологическая роль слюны. Слюноотделение, его регуляция
101. Физиологическая роль печени, участие желчи в пищеварении. Факторы стимулирующие секрецию желчи,
106. Физиология щитовидной и околощитовидной желез
107. Физиология надпочечников. Роль гормонов коры и мозгового вещества в регуляции функции организма
Физиологическая характеристика обонятельной сенсорной системы. Механизмы восприятия запахов
7. Проверить, обладает ли вещество конкурентным действием, можно искусственным введением ацетилхолина. (?)
8. На основании проведенного анализа остается получить результат: данное вещество воздействует на холинорецепторы – курареподобные вещества блокируют их, нарушая открытие Na+каналов на постсинаптической мембране.
№45. При раздражении нерва, иннервирующего мышцу, была зарегистрирована гиперполяризация постсинаптической мембраны. К какому виду относится раздражаемая мышца?
1. Описанное изменение заряда потенциала соответствует утомлению мышцы.
2. Следовательно, синапсы, возбуждаемые при раздражении данного нерва – тормозные.
3. Нервные импульсы, приходящие в варикозы аксонов, высвобождают тормозной медиатор. Воздействуя на постсинаптическую мембрану, тормозной медиатор взаимодействует с хемовозбудимыми каналами, обладающими преимущественной проницаемостью для ионов К+. Выходящий поток калия через эти каналы вызывает гиперполяризацию постсинаптической мембраны.
4. Раздражаемая мышца – гладкая, т.к. именно гладкие мышцы могут возбуждаться и тормозиться благодаря наличию адренергических и холинергических волокон.
№46. В эксперименте на нервно-мышечном препарате лягушки регистрировали биопотенциалы и сокращение мышечного волокна при увеличении частоты его раздражения от 20 до 200 стимулов в секунду. Использовали два способа раздражения мышц – прямое и непрямое. При одном способе раздражения увеличение частоты сопровождалось увеличением амплитуды сокращения и частоты биопотенциалов; при другом способе при увеличенной частоте раздражения мышца расслаблялась, биопотенциалы не регистрировались.
Объясните результат эксперимента при условии, что лабильность раздражаемой мышцы составляла 250 имп/с.
1. При непрямом раздражении мышечного волокна раздражают двигательный нерв, возбуждение по которому посредством синапса передается на мышцу.
2. Доказать, что лабильность мышцы 250 имп/с можно, увеличивая частоту ее раздражения от 200 до 300 стимулов/с. На раздражение 250 и меньше стимулов/с мышца будет отвечать соответствующим числом сокращений в единицу времени в точном соответствии с ритмом наносимых раздражений. Если частота раздражения превышает величину лабильности, то возникает ее трансформация.
3. Лабильность структур при непрямом раздражении: самая большая – нерв, меньше – мышца, еще меньше – синапс.
4. При частоте прямого раздражения 20 имп/с и 200 имп/с мышца будет генерировать ПД с соответствующей частотой. При частоте непрямого раздражения 20 имп/с мышца будет генерировать ПД соответствующей частотой, а при частоте 200 имп/с – с меньшей частотой.
5. Явление уменьшения частоты биопотенциалов и амплитуды мышечного сокращения вплоть до прекращения генерации ПД и расслабление мышц при увеличении частоты раздражения – утомление.
6. При длительном раздражении в нервных окончаниях уменьшается запас медиатора, а его синтез не поспевает за расходованием. Накапливающиеся продукты обмена в мышце понижают чувствительность постсинаптической мембраны к ацетилхолину, в результате чего уменьшается величина постсинаптического потенциала. Когда он понижается до критического уровня, в мышечном волокне не возникает возбуждения. Снижается скорость ресинтеза АТФ, необходимого для мышечного сокращения.
№47. Задачей эксперимента на двух мышцах (А и Б) являлось получение суммарного мышечного сокращения. Предварительно было установлено, что длительность периода укорочения мышцы А – 50 мс, мышцы Б – 500 мс.
1. Для получения слитного суммарного сокращения мышцы А и Б необходимо их раздражать с высокой частотой.
2. Для возникновения суммации необходимо, чтобы интервал между раздражением имел определенную длительность: он должен быть длиннее рефрактерного периода, иначе на второе раздражение не будет ответа, и короче всей длительности сократительного ответа, чтобы второе раздражение подействовало на мышцу раньше, чем она успеет расслабиться после первого раздражения. Причем для возникновения слитного суммированного сокращения раздражение должно приходиться в фазу укорочения.
3. Слитное сокращение мышцы А и мышцы Б – гладкий тетанус.
4. Мышца А относится к медленным красным, Б – к быстрым белым.
5. Продолжительность суммарного сокращение мышцы А будет большей, нежели мышцы Б, т.к. медленные мышечные волокна обладают большей выносливостью, чем быстрые.
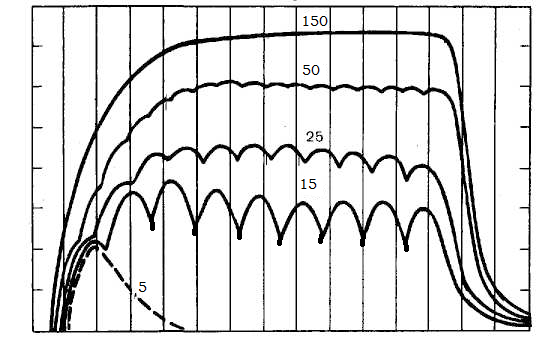
№48. На скелетную мышцу лягушки наносили непрямое раздражение и регистрировали сокращения при частоте 5, 15, 25, 50 и 150 стимулов в секунду. Каковы результаты раздражения?
1. Ответ мышцы на 5 стимулов в секунду – 5 одиночных сокращений, поскольку при воздействии на мышцу непрямых раздражений с большим интервалом, при котором всякое следующее раздражение попадает в период после окончания фазы расслабления.
2. Ответ мышцы на 15 стимулов в секунду – зубчатый тетанус, так как подается много стимулов с интервалом, при котором после первого раздражения сокращение еще не закончилось, а уже возникло новое.
3. Ответ мышцы на 25 стимулов в секунду – зубчатый тетанус, поскольку подается много стимулов с интервалом, при котором после первого раздражения сокращение еще не закончилось, а уже возникло новое.
4. Ответ мышцы на 50 стимулов в секунду – гладкий тетанус, т.к. стимулы наносятся с очень малым интервалом. При сравнении с раздражением 25 стимулов/с последующее раздражение приходится на фазу укорочения, а не расслабления.
5. На непрямое раздражение в 150 имп/с мышца ответит гладким тетанусом, т.к. стимулы наносятся с очень высокой частотой.
6
 .
.№49. Икроножную мышцу и мышцу желудка лягушки помещали в раствор, содержащий ионы Na+, K+, Ca2+ и Cl- в концентрациях, соответствующих их содержанию в тканях. На мышцы наносили прямое надпороговое раздражение и регистрировали биопотенциалы и сокращения.
Как отразилось на результатах эксперимента прибавление к раствору ионов Mg2+, который, как известно, обладает высоким сродством к активным группам кальциевых каналов и, связываясь с ними, блокирует прохождение Ca2+?
1. При блокаде кальциевых каналов нарушается мышечное сокращение.
2. Прибавление ионов магния не повлияло на генерацию ПД икроножной мышцы.
3. Генерация ПД происходит благодаря ионам Na+, К+ и не зависит от ионов Ca2+.
4. Блокада кальциевых каналов повлияла на сокращение скелетной мышцы – его осуществление стало невозможным.
5. Ионы Ca2+ необходимы для сокращения мышцы: они присоединяются к тропонину, и тот прекращает блокировать взаимодействие актина с миозином.
6. Прибавление ионов магния повлияло на генерацию ПД мышцей желудка во время ее раздражения – заблокировало ее.
7. Ионная природа ПД гладкой мышцы определяется особенностями каналов мембраны гладкой мышечной клетки. Основную роль в механизме возникновения ПД играют ионы Са2+.
8. При действии ионов Mg2+ сократимость мышцы желудка изменилась – стала отсутствовать.
9. Ионы Ca2+ перестали поступать в цитоплазму миоцитов желудка. Данные ионы должны связываться с кальмодулином, активирующим киназу легкой цепи миозина, которая приводит к гидролизу АТФ и запускает цикл образования поперечных мостиков.
10. Прибавление ионов магния не влияет на генерацию ПД исчерченной мышцы, однако блокирование кальциевых каналов данными ионами нарушает способность мышцы сокращаться. В гладкой мускулатуре блокирование кальциевых каналов ионами Mg2+ негативно воздействует и на ПД, и на сокращение.
№50. Изолированные икроножные мышцы одной и той же лягушки раздражали с частотой 8 имп/с. Одна из мышц была предварительно утомлена, остальные условия проведения эксперимента были одинаковыми. Оказалось, что сокращения одной из мышц были одиночными, другая отвечала суммарными сокращениями.
Какая мышца отвечала на раздражения одиночными, какая – суммарными сокращениями?
1. Неутомленная мышца не может в данных условиях отвечать на раздражение суммарным сокращением, ведь осуществляется воздействие на мышцу раздражений с большим интервалом, при котором всякое следующее раздражение попадает в период после окончания фазы расслабления.
2. Утомленная мышца может в данных условиях отвечать на раздражение суммарным сокращением. При утомлении нарастает латентный период сокращения и удлиняется период расслабления мышцы, следовательно, последующие сокращения могут попадать в фазу расслабления, вызывая суммарное сокращение.
3. Вывод: неутомленная мышца отвечала на раздражения одиночными, а утомленная – суммарными сокращениями.
| № 1 Известно, что ионные каналы мембраны возбудимой клетки регулируют амплитуду мембранных потенциалов. Экспериментально обнаружено, что яд тетродотоксин блокирует натриевые каналы мембраны возбудимой клетки. Вопросы:1. Как изменится при этом потенциал покоя возбудимости клетки?2. Как изменится при этом потенциал действия возбудимой клетки?3. Как при этом изменится распределение ионов на внешней и внутренней стороне клеточной мембраны? 4. Повлияетли тетродотоксии на проведение возбуждения по нервному волокну? 1. Известно, что потенциал покоя создается преимущественно за счет выхода ионов калия по концентрированному градиенту из клетки. При этом натриевые каналы частично открыты, и некоторое количество ионов натрия проходит в клетку, уменьшая потенциал покоя. Следовательно, блокада натриевых каналов тетродотоксином приведет к небольшому увеличению потенциала покоя. 2. При блокаде натриевых каналов становится невозможным возникновение потенциала действия, так как деполяризация клеточной мембраны невозможна. 3. Концентрация ионов натрия на внешней стороне клеточной мембраны незначительно увеличится, так как ионы натрия полностью перестают входить в клетку. 4. Поскольку проницаемость натриевых каналов увеличивается при возникновении потенциала действия, их блокада тетродоксином приведёт к невозможности распространения возбуждения по нервному волокну. | № 2 Известно, что фазы потенциала действия нервного волокна (быстрая деполяризация и реполяризация) возникают вследствие движения ионов натрия и калия вдоль концентрационных градиентов. В эксперименте на нерв подействовали уабаином — веществом, подавляющим активность АТФазы, затем провели длительное ритмическое раздражение нерва. Вопросы:1. Как при этом изменится распределение ионов на внешней и внутренней стороне клеточной мембраны?2. Изменится ли величина потенциала покоя и потенциала действия в обработанном уабаином нервном волокне? 1. Потенциал покоя и потенциал действия возбудимых клеток обусловлены разной концентрацией ионов, в первую очередь, калия и натрия, снаружи и внутри клетки. Разность концентраций ионов поддерживается благодаря калиево-натриевому насосу, работа, которая является энергозависимой и требует АТФазной активности. Следовательно, ингибирование АТФазы приведет к выравниванию концентраций калия и натрия снаружи и внутри нервных волокон в ходе ритмического раздражения нерва. 2. Отсутствие градиента концентрации ионов снаружи и внутри клетки приведет к исчезновению потенциала покоя и полной невозможности возникновения потенциала действия. | № 3 Известно, что градиент концентрации натрия между двумя сторонами клеточной мембраны влияет на величину потенциалов покоя и действия возбудимой клетки. В эксперименте увеличили концентрацию ионов натрия внутри нервной клетки вначале весьма незначительно, затем выровняли концентрацию ионов натрия внутри клетки и в окружающей клетку среде. Вопрос: Как изменится величина потенциалов покоя и действия в двух описанных ситуациях? 1. Небольшое увеличение концентрации положительно заряженных ионов натрия внутри клетки уменьшит разность потенциалов снаружи и внутри клетки, что вызовет небольшую деполяризацию. Разность потенциалов приблизится к критическому уровню деполяризации, следовательно, возбудимость повысится, и потенциалы действия будут возникать легче. 2. При выравнивании концентрации ионов натрия внутри клетки и снаружи клетки потенциал действия не сможет возникнуть, поскольку натрий не сможет входить в клетку в условиях отсутствия градиента концентрации. | |
| | | ||
| | | ||
| № 4 Известно, что препарат этилендиаминтетраамоний (ЭДТА) связывает в живых тканях ионы кальция. В экспериментальных условиях на нервно-мышечном препарате лягушки в область нервно-мышечного синапса ввели ЭДТА. Вопросы: 1. Как изменится процесс проведения возбуждения в синапсе? 2. Как повлияет ЭДТА на синтез ацетилхолина, проницаемость пресинаптической мембраны для медиатора, генерацию постсинаптического потенциала и активность холинэстеразы? При распространении возбуждения нервного волокна в пресинаптическую область увеличивается проницаемость пресинаптической мембраны и ионы кальция входят по градиенту концентрации внутрь волокна. Связываясь с пресинаптическими везикулами, кальций обеспечивает движение везикул в сторону пресинаптической мембраны, что необходимо для высвобождения медиатора в синаптическую щель. 1. Связывание ионов кальция ЭДТА приведет к прекращению высвобождения ацетилхолина в нервно-мышечном синапсе и блокаде проведения через синапс. 2. ЭДТА не повлияет непосредственно на синтез ацетилхолина, проницаемость пресинаптической мембраны для медиатора, генерацию постсинаптического потенциала и активность холинэстеразы. | № 5 Известно, что проведение возбуждения в синапсе состоит из нескольких стадий. В эксперименте воздействие химического вещества на нервно-мышечные синапсы привело к прекращению передачи возбуждения с нерва на скелетную мышцу. При введении в указанную область ацетилхолина проведение возбуждения через синапс не восстановилось. Введение фермента ацетилхолинэстеразы восстановило проведение возбуждения. Вопросы:1. Перечислите возможные механизмы прекращения проведения возбуждения в синапсе?2. Каков механизм действия изучаемого вещества на нервно-мышечный синапс? 1. Возможными причинами прекращения перехода возбуждения с нерва на мышцу в синапсе могут быть: нарушение выделения медиатора ацетилхолина пресинаптической областью; инактивация или блокада холинэргических рецепторов постсинаптической мембраны; ингибирование фермента, разрушающего ацетилхолин. 2. При добавлении ацетилхолина нервно-мышечная передача не восстановилась, следовательно, дело не в недостаточном выделении медиатора. Добавление фермента холинэстеразы, расщепляющего ацетилхолин и освобождающего рецепторы постсинаптической мембраны для взаимодействия со следующими квантами медиатора, восстановило синаптическую передачу. Следовательно, изучаемое вещество является ингибитором холинэстеразы. | № 6 Известно, что гладкие мышцы имеют ряд физиологических особенностей по сравнению со свойствами скелетных мышц. В ходе эксперимента из стенки кишечника и стенки артерии мышечного типа животного было выделено по фрагменту (длиной 2 см и шириной 2 см), содержащему гладкомышечные волокна. Третий фрагмент такого же размера был выделен из скелетной мышцы. Внешне мало отличающиеся друг от друга мышечные фрагменты поместили в камеру с физиологическим раствором, что обеспечивало условия для их жизнедеятельности в течение некоторого времени. Вопросы:1. Как различить принадлежность фрагментов мышечной ткани по их функциональным свойствам?2. По какому функциональному признаку, без применения воздействий, можно идентифицировать принадлежность одного из фрагментов к мышечной ткани кишечника?3. Как с помощью раздражения фрагментов мышечной ткани можно отличить мышечную ткань внутренних органов от скелетной мышцы? 1. При наличии морфологического сходства между тремя фрагментами мышечной ткани, фрагменты гладкомышечной ткани отличаются автоматизмом, т.е. способностью к спонтанной генерации потенциалов действия и к сокращению. 2. Автоматия хорошо выражена у гладких мышц стенок полых органов, в частности, кишечника, и нехарактерна для гладких мышц стенок кровеносных сосудов. 3. Гладкие мышцы в отличие от скелетных мышц обладают высокой чувствительностью к биологически активным веществам. Подведение раствора ацетилхолина вызовет ритмические сокращения мышечного фрагмента кишечника. Раствор адреналина вызовет спастическое сокращение фрагмента скелетной мышцы. Сокращение фрагмента скелетной мышцы можно вызвать электрическим раздражением. | |
| | | ||
| | | ||
| № 7 Известно, что одним из основных свойств возбудимых тканей является возбудимость. Экспериментально сравнивали возбудимость нервной и мышечной ткани до и после длительного прямого и непрямого раздражения мышцы. Было установлено, что исходно возбудимость одной ткани выше, чем второй. Кроме того, было зафиксировано изменение возбудимости нерва и мышцы после длительного раздражения. Вопросы:1. Как определялась возбудимость нерва и мышцы?2. Какая ткань и почему имела большую возбудимость?3. Как изменилась возбудимость нерва и мышцы после длительного прямого и непрямого раздражения мышцы?4. Какие параметры характеризуют величину возбудимости ткани? 1. На нервно-мышечном препарате лягушки сначала раздражали нерв и затем мышцу одиночными электрическими импульсами. Силу раздражения постепенно увеличивали до появления первого мышечного сокращения. Таким образом, определяли порог раздражения нерва и мышцы. 2. Нервная ткань имела большую возбудимость, так как ее порог раздражения был ниже по сравнению с мышечной тканью. 3. Возбудимость понизится вследствие уменьшения соотношения ионов на клеточной мембране при длительном раздражении нерва и мышцы. 4. Величина возбудимости ткани характеризуется порогом раздражения, реобазой, хронаксией, скоростью аккомодации. | № 8 Известно, что возбудимость является одним из основных свойств нервной и мышечной тканей. Экспериментально было установлено, что после нанесения на изолированный нерв и полоску миокарда надпорогового раздражения возникали потенциалы действия, в ходе которых происходило изменение возбудимости. Вопросы: 1. Какой метод регистрации использовали для изучения потенциалов действия в нерве и полоске миокарда?2. Как изучали изменение возбудимости в различные фазы потенциалов действия?3. Как изменяется возбудимость при возникновении потенциала действия? 1. Биопотенциалы регистрировали с помощью микроэлектродной техники. 2. Изменения возбудимости изучали с помощью измерений порога раздражения в различные фазы потенциалов действия. 3. Во время предспайка возбудимость повышается. При пике потенциала действия возникает абсолютный рефрактерный период. При реполяризации возникает относительный рефрактерный период. При отрицательном следовом потенциале наблюдается фаза экзальтации. При положительном следовом потенциале возникает фаза субнормальной возбудимости. | № 9 Известно, что вокруг клеточных мембран возбудимых тканей существует неравномерное распределение ионов. Экспериментально увеличивали градиент концентрации снаружи и внутри возбудимой клетки раздельно для ионов Na, К, Сl, и Са. Вопросы:Как изменится величина потенциала покоя и потенциала действия при увеличении градиента концентрации раздельно для ионов: 1) Na; 2) К; 3) Сl; 4) Сa? 1. При увеличении градиента концентрации Na величина потенциал покоя снизится, величина потенциала действия увеличится. 2. При увеличении градиента концентрации K величина потенциал покоя возрастет, величина потенциала действия не изменится. 3. При увеличении градиента концентрации Cl величина потенциал покоя возрастет, потенциала действия не изменится. 4. При увеличении градиента концентрации Са величина потенциал покоя снизится, величина потенциала действия не изменится. | |
| | | ||
| | | ||
| № 10 Известно, что процесс возбуждения нервной и мышечной ткани характеризуется изменением ионной проницаемости клеточной мембраны. Экспериментально проводили избирательную блокаду мембранной проницаемости для ионов Na, К, Cl и Са в нервных волокнах и мышечных волокнах скелетной, гладкой и миокардиальной мышц. Вопрос: Как после этого изменится величина потенциалов покоя и действия в этих волокнах? 1. При блокаде мембранной проницаемости для Na величина потенциала покоя повысится, величина потенциала действия понизится. 2. При блокаде мембранной проницаемости для K величина потенциала покоя понизится, величина потенциала действия не изменится. 3. При блокаде мембранной проницаемости для K и для Cl величина потенциала покоя понизится, величина потенциала действия не изменится. 4. При блокаде мембранной проницаемости для K и для Ca величина потенциала покоя повысится, величина потенциала действия не изменится. В миокардиальных и гладкомышечных волокнах длительность потенциалов действия уменьшится в связи с ускорением реполяризации. | № 11 Известно, что суммация одиночных мышечных сокращений является одним из основных свойств мышечной ткани. В экспериментальных условиях изучали способность скелетной мышцы, отрезка кишки и сердца к суммации с помощью нанесения на эти органы двух последовательных раздражений. Вопросы:1. Какие условия надо выполнить, чтобы добиться суммации одиночных сокращений?2. Почему при суммации одиночных сокращений увеличивается амплитуда сокращения?3. Какие виды мышц не способны к суммации одиночных сокращений, и почему это происходит?4. При каких условиях повторяющиеся ритмические раздражения вызывают зубчатый тетанус, гладкий тетанус, оптимум и пессимум скелетной мышцы? 1. Для суммации одиночных сокращений необходимо соблюдение двух условий. Повторное раздражение должно наноситься во время первого одиночного сокращения и не должно попасть в период рефрактерности первого возбуждения. 2. Увеличение амплитуды сокращения при суммации двух одиночных сокращений связано с началом второго сокращения на фоне уже частично сократившейся мышцы. 3. Скелетная мышца способна к суммации одиночных сокращений. Длительность возбуждения и рефрактерного периода в скелетной мышце соответствует латентному периоду ее одиночного сокращения. Если повторное возбуждение попадает во время укорочения или расслабления первого одиночного сокращения, то второе сокращение суммируется с первым. В гладкой мышце длительность возбуждения и рефрактерный период соответствуют латентному периоду и фазе укорочения ее одиночного сокращения. Поэтому суммация двух сокращений может произойти в случае попадания второго раздражения в фазу расслабления гладкой мышцы. В сердце длительность возбуждения и рефрактерный период равны длительности одиночного сокращения. Поэтому суммация одиночных сокращений в миокарде не происходит. 4. Зубчатый тетанус возникает, когда каждое последующее раздражение попадает в фазу расслабления предыдущего одиночного сокращения. Гладкий тетанус возникает, когда каждое последующее раздражение попадает в фазу укорочения предыдущего одиночного сокращения. Оптимум возникает, когда каждое последующее раздражение попадает в фазу экзальтации предыдущего возбуждения. Пессимум возникает, когда каждое последующее раздражение попадает в абсолютный рефрактерный период предыдущего возбуждения. | № 12 Известно, что поддержание постоянства ионного состава в возбудимых тканях необходимо для их нормального функционирования. Отсутствие солей (и в частности — солей кальция) в питьевой воде у населения отдельного района России привело к нарушению функций скелетной мускулатуры. Обследование населения показало, что даже практически здоровые люди жаловались на повышенную мышечную утомляемость и недостаточную физическую силу. Вопросы:1. Какова роль ионов кальция в механизме мышечного сокращения?2. Почему недостаток кальция в организме сопровождается повышенной физической утомляемостью и недостаточной физической силой у людей? 3. Какие обследования целесообразности для изучения функций скелетной мускулатуры и миокарда? 1. В расслабленном мышечном волокне белок тропонин препятствует взаимодействию головок выростов миозина с актином. Мышечное сокращение начинается с выхода ионов кальция из Т-систем и эндоплазматического ретикулума. Ионы кальция соединяются с тропонином, и он сдвигается в сторону. Головки выростов миозина вступают в контакт с актином, обеспечивая скольжение нитей актина вдоль миозина. Длина саркомеров миофибрилл уменьшается. Длина всей мышцы уменьшается. 2. Количество связанных с тропонином ионов кальция определяет количество поперечных мостиков между нитями актина и миозина и, следовательно, силу и длительность сокращения. Поэтому недостаток в организме ионов кальция у людей приводит к уменьшению мышечной силы и повышению физической утомляемости. 3. Для изучения силы и выносливости скелетной мускулатуры необходимо исследовать динамометрию. Для изучения выносливости сердца необходимо провести исследование функций сердца с физическими нагрузками. | |
| | | ||
| | |
| № 13 Известно, что ацетилхолин является одним из основных медиаторов нервной системы. Входе обследования испытуемого было установлено, что блокатор ацетилхолинергической передачи возбуждения в синапсах атропин вызвал расширение зрачка, увеличение частоты и силы сердечных сокращений, уменьшение перистальтики желудочно-кишечного тракта. При этом не изменилась сократительная функция скелетной мускулатуры. Вопросы:1. На какие постсинаптические рецепторы действует ацетилхолин при выделении его в синаптическую щель?2. Объясните возможные причины различного действия атропина в нервно-мышечных синапсах соматической нервной системы и в синапсах вегетативной нервной системы на внутренних органах. 1. Медиатор ацетилхолин действует на два вида постсинаптических рецепторов: М- и Н-холинорецепторы. М-холинорецепторы находятся в нейроорганных синапсах парасимпатической нервной системы. Н-холинорецепторы находятся в нервно-мышечных синапсах и вегетативных ганглиях. 2. Атропин блокирует только М-холинорецепторы. Поэтому блокируется действие парасимпатической нервной системы, а соматическая регуляция скелетной мускулатуры не нарушается. | № 14 Известно, что суммарный потенциал действия нерва складывается из потенциалов действия одиночных нервных волокон, входящих в нерв. Экспериментально исследовали суммарный потенциал действия изолированного седалищного нерва, выделенного из крупного животного. Раздражение наносили на проксимальный конец нерва. На дистальном конце нерва суммарный потенциал имел сложную форму и состоял из нескольких пиков и волн. Амплитуда его значительно уменьшилась. Вопросы:1. С чем связано изменение формы суммарного потенциала действия по ходу проведения возбуждения в нерве?2. Как диаметр нервного волокна и наличие миелина влияют на скорость проведения возбуждения? 3. Почему происходит уменьшение амплитуды суммарного потенциала? 4. Работает ли при этом закон «Всё или ничего»? 1. Расслоение суммарного потенциала действия на отдельные волны связано с различной скоростью проведения возбуждения в волокнах, образующих нерв. 2. Скорость проведения возбуждения больше в миелиновых волокнах с большим диаметром. 3. Амплитуда суммарного потенциала уменьшается вследствие уменьшения количества нервных волокон на дистальном конце нерва, что связано с ответвлениями от нерва нервных волокон по ходу его длины. 4. Закон “все или ничего” при этом не работает. Суммарный потенциал действия нерва прямо зависит от количества волокон входящих в нерв. Потенциал действия каждого волокна, входящего в нерв, подчиняется закону “все или ничего”. | № 15 Известно, что утомляемость является одним из основных свойств возбудимой ткани. В эксперименте на нервно-мышечном препарате лягушки проводили ритмическое непрямое раздражение мышцы. Вопросы:1. Как изменится амплитуда мышечного сокращения после длительного раздражения нерва?2. Как изменится амплитуда и частота потенциалов действия в нервных волокнах при развитии утомления в мышце? 1. При длительном раздражении амплитуда мышечных сокращений уменьшается за счет процесса утомления мышцы. 2. Амплитуда и частота потенциалов действия нервных волокон не изменится, так как нервы подчиняются закону относительной неутомляемости. | |
| | | ||
| | | ||
| № 16 Известно, что лабильность, или функциональная подвижность, является важной характеристикой возбудимых тканей. Экспериментально была исследована лабильность возбудимых тканей лягушки: седалищного нерва, состоящего из миелиновых волокон, одного из симпатических нервов, состоящего из безмиелиновых волокон, нервно-мышечных синапсов и икроножной мышцы. Вопросы: 1. Как определяли лабильность возбудимых образований? 2. Какой параметр используют как меру лабильности?3. У каких исследованных образований лабильность выше или ниже и почему? 4. Как лабильность может характеризовать возбудимость исследуемых образований? 1. Лабильность определяют с помощью нанесения ритмических раздражений с увеличивающейся частотой и регистрацией процессов возбуждения или сокращения в исследуемой возбудимой ткани. 2. Мерой лабильности является максимальная частота раздражения, которую возбудимая ткань может воспроизвести без трансформации ритма. 3. Мера лабильности обратно пропорциональна длительности рефрактерного периода. Миелиновые нервные волокна обладают наибольшей лабильностью. Безмиелиновые нервные волокна имеют меньшую лабильность. Лабильность мышечных волокон еще меньше. Наименьшей лабильностью обладают синапсы, что связано с задержкой проведения возбуждения в синапсах. 4. Лабильность является одной из косвенных характеристик возбудимости. Чем больше лабильность, тем больше возбудимость. | № 17 В эксперименте на животном при действии светового, звукового или тактильного раздражителей в коре головного мозга возникают вызванные электрические потенциалы. Вопросы:1. По каким путям импульсы от соответствующих рецепторов поступают в кору головного мозга?2. В каких отделах коры наблюдаются вызванные электрические потенциалы: а) при световых воздействиях; б) при акустических воздействиях; в) при тактильных воздействиях? 1. Возбуждения, возникшие в фоторецепторах сетчатки в ответ на действие светового стимула, поступают по зрительному нерву к верхним буграм четверохолмия, наружным коленчатым телам и далее к коре головного мозга. Возбуждения от механорецепторов внутреннего уха и клеток спирального ганглия поступают по слуховой части вестибулокохлеарного нерва в нижние бугры четверохолмия, внутренние коленчатые тела и далее в кору головного мозга. Возбуждения от тактильных рецепторов кожи поступают по задним канатикам спинного мозга к ядрам задних столбов и дальше в специфические ядра таламуса и в кору головного мозга. 2. а — в первичной и вторичной зрительной затылочной коре на свет; б — в первичной и вторичной слуховой области височной коры на звук; в — в первой и второй соматосенсорной области постцентральной извилины на тактильное раздражение. | № 18 У собаки на фоне механического раздувания специальным баллончиком прямой кишки осуществляют электрическое раздражение участка сенсомоторной коры. Вопросы:1. Какой эффект при этом наблюдается?2. Как объяснить подобный эффект?3. Какое свойство нервных центров лежит в основе данного эффекта? 1. У животного возникает дефекация. 2. Причиной подобной реакции является электрическое раздражение коры, которое усиливает возбуждение структур мозга, возникшее при механическом раздражении кишечника. 3. Способность к формированию на основе физиологической потребности соответствующего доминирующего возбуждения. | |
| | | ||
| | | ||
| № 19 В эксперименте у собаки проведено полное удаление мозжечка. Вопросы:1. Какие нарушения поведения наблюдаются при этом у животного?2. Какие функции выполняет мозжечок?3. С какими структурами мозга связан мозжечок, и какое функциональное значение имеют эти связи? 1. Абазия, атония, атаксия, астазия, астения. 2. Формирование программы движения, контроль выполнения движения, коррекция нарушения движения, вегетативное обеспечение движения. 3. Через верхние ножки мозжечок получает информацию из ассоциативных областей коры о цели действия, через нижние (из спинного мозга) — о состоянии опорно-двигательного аппарата (положение тела и конечностей), через средние — команды от мозжечка к ядрам ствола мозга и к двигательной коре больших полушарий. | № 20 Для изучения деятельности спинальных нервных центров у лягушки последовательно перерезают нервные корешки, связывающие спинной мозг с периферией. Вопросы:1. Какие функции выполняют передние и задние корешки спинного мозга?2. Какой эффект наблюдается при перерезке у лягушки всех задних корешков с левой стороны?3. Какой эффект наблюдается при перерезке всех передних корешков правой стороны? 1. Передние корешки являются эфферентными двигательными, а задние — афферентными чувствительными. 2. После перерезки у лягушки всех дорсальных корешков левой стороны исчезает сгибательный тонус левых конечностей. 3. При перерезке у лягушки всех вентральных корешков правой стороны исчезают движения правых конечностей. | № 21 Как известно, в деятельности головного мозга имеет место процесс торможения. В процессе рассматривания сложного изображения или прослушивания музыкального фрагмента испытуемый выделяет их световые, цветовые и звуковые характеристики. Вопросы:1. Дайте определение центрального торможения. 2. Какие виды центрального торможения вам известны? 3. Какой вид центрального торможения лежит в основе улучшения различий частоты звуков, выделения контуров изображения, дифференциации соседних точек прикосновения на коже? 1. Физиологический процесс, возникающий в центральной нервной системе на основе возбуждения и приводящий либо к уменьшению, либо к его полному прекращению. 2. Возвратное, реципрокное, латеральное торможение. 3. Латеральное торможение. | |
| № 22 При раздражении слабым электрическим током коры головного мозга животного в эксперименте наблюдаются сокращения отдельных мышц туловища и конечностей. Вопросы:1. Какие отделы коры головного мозга при этом раздражаются?2. Какой объем движений (отдельные мышечные волокна, целые мышцы, движения в суставе) наблюдается в этих случаях?3. На что указывают размеры представительства конечностей в коре головного мозга? 1. Моторные области прецентральной извилины. 2. Как отдельные мышцы, так и группы мышц, формирующие движение в суставе. 3. Размеры представительства движений в моторной коре объясняются числом моносинаптических связей между аксонами пирамидных нейронов коры и мотонейронами, иннервирующими мышцы головы, туловища, конечностей. | № 23 В эксперименте на обезьяне регистрируется импульсная активность нервов, несущих информацию от проприоцепторов мышцы-сгибателя и рецепторов сумки локтевого сустава. Вопросы:1. Какие рецепторы мышц и суставов вы знаете?2. Какую информацию посылают данные рецепторы при сгибании и разгибании конечности в локтевом суставе? 3. Сохранятся ли движения конечности в суставе после перерезки указанных нервов? 1. Нервы мышечных веретен, рецепторы сухожильного органа Гольджи, рецепторы суставной сумки. 2. Рецепторы мышечных веретен трехглавой мышцы посылают информацию о степени, скорости и ускорении ее растяжения; рецепторы сухожильного органа Гольджи — о величине мышечного напряжения; суставные рецепторы — о величине и скорости изменения угла между предплечьем и плечом. 3. Сохранятся. | № 24 При разрушении в эксперименте некоторых структур головного мозга у обезьяны возникает синдром Клювера-Бьюси. Вопросы: 1. Какие структуры при этом разрушаются? 2. Какие изменения поведения в этом случае имеют место? 3. Какие функции выполняет отдел головного мозга, в состав которого входят разрушенные структуры. 1. Височная кора и гиппокамп. 2. Гиперсексуальность, оральные реакции, агрессивность. 3. Лимбическая система участвует в обеспечении механизмов памяти, мотиваций, эмоций. | |
| | | ||
| № 25 Во время нейрохирургической операции у пациента при раздражении коры слабым электрическим током возникли ощущения давления, прикосновения или тепла на коже противоположной стороны. Вопросы: 1. Какие отделы коры раздражались? 2. Различаются ли представительства участков кожной поверхности туловища и конечностей в коре (“сенсорный гомункулюс”)? 3. Нарисуйте пути проведения импульсов от кожных рецепторов в кору больших полушарий. 1. Задняя постцентральная извилина. 2. Структура представительства различных видов чувствительности в соматосенсорной коре отражает различие в числе рецепторов, расположенных в коже головы, туловища и конечностей. 3. Дается рисунок проведения импульсов от кожных рецепторов до коры. | № 26 При поперечной перерезке ствола мозга у экспериментального животного наблюдается состояние децеребрационной ригидности. Вопросы:1. В чем это состояние проявляется?2. Между какими структурами нужно сделать перерезку для получения указанного состояния?3. Какие механизмы лежат в основе децеребрационной ригидности? 1. Состояние децеребрационной ригидности проявляется повышением тонуса мышц-разгибателей туловища и конечностей. 2. Состояние децеребрационной ригидности достигается поперечным разрезом мозга ниже красных ядер. 3. Перерезка приводит к устранению тормозного влияния красных ядер на сегментарный аппарат спинного мозга. | № 27 У децеребрированной кошки при пассивном повороте головы в правую сторону увеличивается тонус мышц-разгибателей обеих правых конечностей; при повороте головы влево — левых конечностей. Вопросы:1. Какие причины лежат в основе данного явления?2. Какие виды тонических рефлексов вы знаете?3. В каких отделах центральной нервной системы находятся центральные звенья тонических рефлексов? 1. При повороте головы децеребрированного животного с сохраненными лабиринтами в одну сторону увеличивается активность вестибулоспинальной системы, повышающей тонус мышц-разгибателей той же стороны. 2. Тонические рефлексы бывают статические и статокинетические. Статические подразделяются на рефлексы положения и рефлексы выпрямления. Статокинетические подразделяются на рефлексы в случаях прямолинейного движения тела и рефлексы при круговых движениях. 3. Центры тонических рефлексов расположены в ядрах моста и продолговатого мозга. | |
| | | | |
| № 28 В эксперименте на кролике электрическое раздражение гипоталамуса, таламуса и ретикулярной формации вызывает характерные изменения электрической активности коры головного мозга. Вопросы: 1. В чём эти изменения проявляются? 2. В чем причина этих изменений? 3. Какие основные функции обеспечиваются данными структурами головного мозга? 1. При раздражении гипоталамуса наблюдается ограниченная реакция активации ЭЭГ в передних отделах коры головного мозга. 2. При раздражении таламуса — в первичных сенсорных проекционных зонах. 3. При раздражении ретикулярной системы — генерализованная активация во всех отделах коры. В естественных условиях активация гипоталамуса отражает возникновение мотивационного возбуждения, таламуса — приход возбуждений от рецепторов органов чувств, ретикулярной формации — восходящие активирующие влияния, имеющие мотивационную окраску. | № 29 У животного в эксперименте проведена перерезка спинного мозга. Вопросы:1. Какие симптомы имеют место у животного после исчезновения спинного шока? 2. Каковы механизмы появления установленных симптомов?3. Какие функции спинного мозга вам известны? 1. После травмы по прошествии спинального шока утрачиваются произвольные движения конечностей, наблюдается повышение тонуса скелетной мускулатуры и выпадение всех видов чувствительности туловища и конечностей. 2. Полное прекращение связей спинного мозга с вышерасположенными отделами головного мозга. 3. Сегментарно-рефлекторная функция, проводниковая функция, функция автоматии центров спинного мозга. | № 30 В неврологическое отделение поступил больной с высоким обрывом спинного мозга, наступившим в результате автомобильной аварии. Вопросы: 1. Какие симптомы имеют место у пострадавшего? 2. Каковы механизмы появления установленных симптомов? 3. Какие функции спинного мозга Вам известны? 1. после травмы по прошествии спинального шока утрачиваются произвольные движения конечностей, повышение мышечного тонуса и выпадение всех видов чувствительности туловища и конечностей. 2. Полное прекращение связей спинного мозга с вышерасположенными отделами головного мозга. 3. Сегментарно-рефлекторная функция, проводная функция, функция центров автоматии спинного мозга. | |
| | | | |
| № 31 В клинику поступил больной с кровоизлиянием в структуры продолговатого мозга. Вопросы:1. Какие основные центры продолговатого мозга вам известны?2. Какие симптомы наблюдаются при поражении продолговатого мозга?3. В чем заключается опасность поражения продолговатого мозга? 1. Сердечно-сосудистый центр; дыхательный центр; центры защитных реакций: рвотный, чихательный, кашлевой; центры некоторых тонических рефлексов. 2. Нарушения сердечно-сосудистой и дыхательной функций. 3. Остановка сердца, коллапс сосудов, остановка дыхания. | № 32 При действии раздражителей на экстеро или интерорецепторы импульсы от последних поступают в центральные отделы анализаторов. Вопрос: Какими механизмами могут распространяться поступление возбуждения в центральной нервной системе? Иррадиация, суммация, конвергенция, мультипликация, промонгирование, оккмозия. | № 33 В опыте И.М.Сеченова для демонстрации центрального торможения предварительно осуществляют глубокий поперечный разрез на уровне зрительных бугров. Вопрос: Какая цель достигается данной операцией? Для исключения влияния центров коры больших полушарий. | |
| | | | |
| № 34 В каком компоненте рефлекторной дуги, как и в каких условиях протекают процессы, приводящие к изменению времени рефлекторной реакции? В центральном компоненте, а также на уровне рецептора и афферентного волокна. | № 35 Для изучения деятельности нервных спинальных центров животных выполняют операцию перерезки спинного мозга или нервов, ведущих к этому центру. Вопрос: Какой эффект наблюдается у лягушек при перерезке всех задних корешков с левой стороны или всех передних корешков правой стороны тела? При перерезке задних корешков слева исчезает чувствительность и снижается двигательная активность. При перерезке передних корешков исчезает двигательная активность. | № 36 В процессе обучения игре на музыкальных инструментах или печатанью на компьютере приобретается лёгкость и автоматизм двигательного навыка. Вопрос: Какими свойствами нервных центров можно объяснить эти явления? Эти явления можно объяснить свойством пластичности доминанта, тонусом нервного центра. | |
| | | ||
| | |